Խոլեստերինի սինթեզի կարգավորումը
Խոլեստերինի սինթեզի կարգավորումը `դրա հիմնական ֆերմենտը (HMG-CoA reductase) իրականացվում է տարբեր եղանակներով:
HMG ռեդուկտազի ֆոսֆորիլացիա / dephosphorylation: Ինսուլին / գլյուկագոն հարաբերակցության բարձրացումով, այս ֆերմենտը ֆոսֆորիլացնում է և անցնում ակտիվ վիճակի: Ինսուլինի գործողությունը իրականացվում է 2 ֆերմենտների միջոցով:
HMG-CoA reductase kinase phosphatase, որը kinase- ը վերածում է անգործուն dephosphorylated պետության:
Phosphotase HMG-CoA reductase- ը այն վերածելով dephosphorylated ակտիվ պետության: Այս ռեակցիաների արդյունքը HMG-CoA ռեդուկտազի դեֆոսֆորիլացված ակտիվ ձևի ձևավորումն է:
Հետևաբար, կլանման ժամանակահատվածում խոլեստերինը մեծանում է: Այս ժամանակահատվածում ավելանում է խոլեստերինի `ացետիլ - CoA- ի սինթեզի համար նախնական սուբստրատի առկայությունը (ածխաջրեր և ճարպեր պարունակող սնունդ ուտելու արդյունքում, քանի որ CoA- ի ացետիլը ձևավորվում է գլյուկոզի և ճարպաթթուների տրոհման ժամանակ):
Հետցնցումային վիճակում գլյուկագոնը A պրոտեինոգենազի միջոցով խթանում է HMG - CoA- ի ֆոսֆորիլացումը `ռեդուկտազը, այն վերածելով ոչ ակտիվ վիճակի: Այս գործողությունը ուժեղանում է նրանով, որ միևնույն ժամանակ գլյուկագոնը խթանում է HMG-CoA reductase ֆոսֆոտազի ֆոսֆորիլացումը և անգործությունը, դրանով իսկ HMG-CoA- ի ռեդուկտազը պահելով ֆոսֆորիլացված ոչ ակտիվ վիճակում: Արդյունքում, հետբաբսորման ժամանակահատվածում և ծոմապահության ժամանակ խոլեստերինի սինթեզը արգելվում է էնդոգեն սինթեզով: Եթե սննդի մեջ խոլեստերին պարունակությունը բերվել է 2% -ի, ապա էնդոգեն խոլեստերինի սինթեզը կտրուկ նվազել է: Բայց խոլեստերինի սինթեզի ամբողջական դադարեցումը տեղի չի ունենում:
Խոլեստերինի բիոսինթեզի արգելակման աստիճանը սննդից եկող խոլեստերինի ազդեցության տակ տատանվում է անձից այլ անձի: Սա ցույց է տալիս խոլեստերինի ձևավորման գործընթացների անհատականությունը: Նվազեցնելով խոլեստերինի սինթեզի ինտենսիվությունը, հնարավոր է նվազեցնել նրա կոնցենտրացիան արյան մեջ:
Եթե մի կողմից մարմնում խոլեստերին ընդունողկազմի և մի կողմից մարմնում դրա սինթեզի միջև հավասարակշռությունը խախտվում է, մյուս կողմից `լեղաթթուների և խոլեստերինի արտանետումների միջև, խախտվում է, հյուսվածքներում խոլեստերինի կոնցենտրացիան փոխվում է: Ամենալուրջ հետևանքները կապված են արյան խոլեստերինի համակենտրոնացման բարձրացման հետ (հիպերխոլեստերեմիա), մինչդեռ մեծանում է աթերոսկլերոզի և խոլելիտիասի զարգացման հավանականությունը:
Ընտանեկան հիպերխոլեստերեմիա (HCS) - այս ձևը շատ ավելի տարածված է `200 մարդու մոտ 1 հիվանդ: HCS- ում ժառանգված թերությունը բջիջների կողմից LDL- ի կլանման խախտում է, և, հետևաբար, LDL կատաբոլիզմի մակարդակի նվազում: Արդյունքում, արյան մեջ LDL- ի կոնցենտրացիան բարձրանում է, ինչպես նաև խոլեստերինը, քանի որ LDL- ում դրա մեծ մասը կա: Հետևաբար, HCS- ով բնորոշ է հյուսվածքներում խոլեստերինի պահպանումը, մասնավորապես մաշկում (քսանտոմա), զարկերակների պատերին:
HMG-CoA ռեդուկտազի սինթեզի խոչընդոտում
Խոլեստերինի նյութափոխանակության ուղու վերջնական արտադրանքը: Այն նվազեցնում է HMG-CoA reductase գենի արտագրման տեմպը ՝ դրանով իսկ խանգարելով սեփական սինթեզին: Լյարդը ակտիվորեն սինթեզում է լեղու թթուները խոլեստերինից, և, հետևաբար, լեղաթթուները խանգարում են HMG-CoA reductase գենի գործունեությունը: Քանի որ HMG-CoA ռեդուկտազը գոյություն ունի մոտ 3-ի սինթեզից հետո, այս խոլեստերինի ֆերմենտի սինթեզի արգելակումը արդյունավետ կարգավորում է:
Չե՞ք գտել այն, ինչ փնտրում եք: Օգտագործեք որոնումը.
Խոլեստերինի էսթերի փոխանակում
Խոլեստերինի ֆոնդը պարունակում է անվճար խոլեստերին և խոլեստերինի էթերներ, որոնք հանդիպում են ինչպես բջիջներում, այնպես էլ արյան լիպոպրոտեիններում:
II մաս Մետաբոլիզմ և էներգիա
Խցերում խոլեստերինի էստերիֆիկացումը տեղի է ունենում ացիլ-CoA- խոլեստերին-acyltransferase (AChAT) գործողությամբ:
Acyl-CoA + խոլեստերին - * HS-KoA + Acylcholesterol
Մարդկային բջիջներում հիմնականում ձևավորվում է լինոլեյխոլեստերին: Ի տարբերություն անվճար խոլեստերինի, բջիջների մեմբրաններում դրա էսթերները պարունակվում են շատ փոքր քանակությամբ և հիմնականում հայտնաբերվում են ցիտոզոլում ՝ որպես լիպիդային կաթիլների մի մաս: Էսթերների ձևավորումը մի կողմից կարելի է համարել որպես թաղանթներից ավելցուկային խոլեստերին հեռացնելու մեխանիզմ, իսկ մյուս կողմից ՝ որպես բջիջում խոլեստերին խտացնելու մեխանիզմ: Ռեսուրսների մոբիլիզացումը տեղի է ունենում էստերազային ֆերմենտների մասնակցությամբ, որոնք հիդրոլիզացնում են խոլեստերինի եթերները.
Acylcholesterol + H 2O - * ճարպաթթու + խոլեստերին
Էստերի սինթեզը և հիդրոլիզացումը տեղի են ունենում բազմաթիվ բջիջներում, բայց հատկապես ակտիվ է վերերիկամային ծառի կեղևի բջիջներում. Այդ բջիջներում մինչև խոլեստերինի մինչև 80% -ը ներկայացված են էստերով, իսկ մյուս բջիջներում սովորաբար 20% -ից պակաս է:
Արյան լիպոնտրոտիններում էսթերների ձևավորումը տեղի է ունենում լեցիտին-խոլեստերինի ացիլtransferase- ի (LHAT) մասնակցությամբ, որը կատալիզացնում է լազիտինի i- դիրքերից խոլեստերինի ացիլային մնացորդի տեղափոխումը (Նկար 10.31): LHAT- ը ձևավորվում է լյարդի մեջ, որը արտազատվում է արյան մեջ և կցվում է լիպոպրոտեինների: Տարբեր լիպոպրոտեինների համար էսերիֆիկացման մակարդակը զգալիորեն տարբերվում է և կախված է ապոլիպոպրոտեինների առկայությունից, որոնք ակտիվացնում են LHAT (հիմնականում `AP-AT, ինչպես նաև CI) կամ խանգարում (C-II) այս ֆերմենտը: HDL- ի առավել ակտիվ LHAT- ը, որում AP-AT պետք է Olee 2/3 բոլոր սպիտակուցների: Ամենամեծ coli գերազանցությամբ ձեւավորված եթերների Oleic եւ linoleic թթուներ. Մյուս լիպոպրոտեինները ester ձեւավորումը տեղի է ունենում դանդաղ տեմպերով, քան HDL.
Նկ. 10.31. Խոլեստերինի էսթերների ձևավորում LHAT գործողության ներքո
LHAT- ը տեղայնացված է HDL- ի մակերևութային շերտում և ֆոսֆոլիպիդային միաձուլման մեջ օգտագործում է խոլեստերին ՝ որպես ենթաշերտ: Այստեղ ձևավորված խոլեստերինի էթերները, ամբողջական հիդրոֆոբիկության պատճառով, վատ են պահպանվում
Գլուխ 10. Մետաբոլիզմը և լիպիդային ֆունկցիան
ֆոսֆոլիպիդային միաձուլիչ և ընկղմված է լիպոպրոտեինների լիպիդային հիմքում: Միևնույն ժամանակ, խոլեստերինի համար տեղ ազատվում է ֆոսֆոլիպիդային միաձուլման մեջ, որը խոլեստերինով կարող է լցվել բջջային թաղանթներից կամ այլ լիպոպրոտեիններից: Այսպիսով, LHAT գործողության արդյունքում HDL- ը խոլեստերինի ծուղակ է թվում:
Bile թթու սինթեզ
Լյարդում խոլեստերինի մի մասը վերածվում է լեղու թթուների: Գալաթթուները կարող են համարվել որպես քոլանաթթվի ածանցյալներ (Նկար 10.32):
Քոլանաթթուն, որպես այդպիսին, մարմնում չի ձևավորվում: Հեպատոցիտներում խոլեստերինն ուղղակիորեն արտադրում է ցենոդեօքսիխոլական և խոլաթթուներ `առաջնային աղու թթուներ (Նկար 10.33, տե՛ս նաև Նկար 10.12):
Խոլեստերինի բիոսինթեզ
Խոլեստերինի բիոսինթեզը տեղի է ունենում էնդոպլազմիկ ցանցաթաղանթում: Մոլեկուլում ածխածնի բոլոր ատոմների աղբյուրը ացետիլ-SCoA- ն է, որն այստեղ գալիս է mitochondria- ից `որպես ցիտրատի մի մաս, ինչպես ճարպաթթուների սինթեզում: Խոլեստերինի կենսոսինթեզը սպառում է 18 ATP մոլեկուլ և 13 NADPH մոլեկուլ:
Խոլեստերինի ձևավորումը տեղի է ունենում ավելի քան 30 ռեակցիա, որը կարող է խմբավորվել մի քանի փուլով:
1. Մեվալոնաթթվի սինթեզ:
Սինթեզի առաջին երկու ռեակցիաները համընկնում են ketogenesis- ի ռեակցիաների հետ, բայց 3-հիդրօքսի-3-մեթիլգլուտարիլ-ScoA- ի սինթեզից հետո ֆերմենտը մտնում է հիդրօքսիմեթիլ-գլուտարիլ-ScoA ռեդուկտազը (HMG-SCOA reductase) ՝ ձևավորելով մեվալոնաթթու:
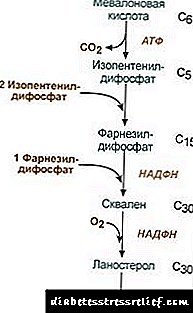 |
Խոլեստերին սինթեզի ռեակցիայի սխեման2. Իզոպենթենիլ դիֆոսֆատի սինթեզ: Այս փուլում ֆոսֆատի երեք մնացորդները կցվում են մեվալոնաթթուին, այնուհետև այն ապարբոքսիլացվում է և ջրազրկվում: 3. Իզոպանտենիլ դիֆոսֆատի երեք մոլեկուլները համատեղելուց հետո սինթեզվում է ֆարնեսիլ դիֆոսֆատը: 4. Squalene- ի սինթեզը տեղի է ունենում այն ժամանակ, երբ երկու ֆարնեսիլ դիֆոսֆատի մնացորդները կապված են: 5. Բարդ ռեակցիաներից հետո գծային սկվալենը ցիկլիզացնում է լանոստերոլը: 6. Մեթիլների ավելցուկային հեռացումը, մոլեկուլի վերականգնումն ու իզոմերիացումը հանգեցնում են խոլեստերինի տեսքի: Հիդրոքսիմեթիլգլուտարիլ-S-CoA ռեդուկտազի գործունեության կարգավորում3. Խոլեստերինի բիոսինթեզի արագությունը նույնպես կախված է համակենտրոնացումից հատուկ կրիչ սպիտակուցապահովելով հիդրոֆոբային միջանկյալ սինթեզի մետաբոլիտների կապը և տեղափոխումը: Կարող եք հարցնել կամ թողնել ձեր կարծիքը: Կարգավորման հիմնական կետը մեվոլոնաթթվի ձևավորման արձագանքն է: 1. Ալլաստերի կարգավորումը: Խոլեստերինը, և լյարդի մեջ - և լեղաթթուները խանգարում են HMG-CoA ռեդուկտազին: 2. HMG-CoA reductase խոլեստերինի սինթեզի ռեպրեսիան: 3. Կարգավորումը HMG-CoA ռեդուկտազի ֆոսֆորիլ-դեֆոսֆորիլացման միջոցով, ակտիվ ոչ ֆոսֆորիլացված ձևով: Գլյուկագոնը դեակտիվացում է առաջացնում, իսկ ինսուլինը առաջացնում է ակտիվացում ռեակցիաների բարդ կասկադի միջոցով: Այսպիսով, խոլեստերինի սինթեզի արագությունը փոխվում է ներծծող և հետընտրական կլանման վիճակների փոփոխությամբ: 4. Լյարդում HMG-CoA ռեդուկտազի սինթեզի արագությունը ենթակա է ակնհայտ Diurnal տատանումների. Առավելագույնը կեսգիշերին և առավոտյան նվազագույն: Խոլեստերինի էսթերի փոխանակում Խցերում խոլեստերինի էսթերացումը տեղի է ունենում, երբ ենթարկվում է acyl-CoA- խոլեստերին-acyltransferase (ԱՀԱՏ). Acyl-CoA + Խոլեստերին ® NS-CoA + Acylcholesterol Խցերում հիմնականում ձևավորվում է լինոլեյլխոլեստերին: Էստերը հայտնաբերվում են հիմնականում ցիտոզոլում `որպես լիպիդային կաթիլների մի մաս: Էսթերների ձևավորումը մի կողմից կարելի է համարել որպես թաղանթներից ավելցուկային խոլեստերին հեռացնելու մեխանիզմ, իսկ մյուս կողմից ՝ որպես բջիջում խոլեստերին խտացնելու մեխանիզմ: Պաշարների մոբիլիզացումը տեղի է ունենում ֆերմենտների մասնակցությամբ էստերազխոլեստերինի էստերների հիդրոլիզացումը. Acylcholesterol + H2O ® ճարպաթթու + խոլեստերին Էստերի սինթեզը և հիդրոլիզացումը հատկապես ակտիվ են վերերիկամային ծառի կեղևի բջիջներում: Արյան լիպոպրոտեիններում ester- ի ձևավորումը տեղի է ունենում մասնակցությամբ լեցիտին-խոլեստերին-acyltransferase (LHAT) ՝ կատալիզացնելով ացիլային մնացորդի տեղափոխումը լեցիտինից խոլեստերին: LHAT- ը ձևավորվում է լյարդի մեջ, գաղտնազերծվում է արյան մեջ և կցվում է լիպոպրոտեինների: Առավել ակտիվ LHAT- ը HDL- ում, որտեղ տեղայնացված է մակերևույթի շերտում: Այստեղ ձևավորված խոլեստերինի էթերները հիդրոֆոբ են և ընկղմվում են լիպիդային միջուկում: Ֆոսֆոլիպիդային միաձուլման մեջ խոլեստերինի համար ազատ տեղ կա, որը կարող է խոլեստերինով լցվել բջջային մեմբրաններից կամ այլ լիպոպրոտեիններից: Այսպիսով, LHAT գործողության արդյունքում HDL- ը խոլեստերինի ծուղակ է թվում: Bile թթու սինթեզ Լյարդում խոլեստերինի մի մասը վերածվում է լեղու թթուների: Ոսկրածուծի թթուները կարող են համարվել որպես քոլանաթթվի ածանցյալներ: Քոլանաթթուն, որպես այդպիսին, մարմնում չի ձևավորվում: Խոլեստերինից հեպատոցիտներում ձևավորվում են առաջնային լեղու թթուներ. ցենոդոքսիկոլիկ և խոլիկ. Աղիքային ֆլորայի ֆերմենտների գործողության միջոցով աղիների մեջ աղիների գաղտնի արտազատումից հետո դրանցից ձևավորվում են երկրորդական լեղու թթուներ. լիտոլոգիական և դեօքսիխոլիկ. Դրանք ներծծվում են աղիքներից, պորտալարի երակային արյունը մտնում է լյարդը, այնուհետև `լեղապարկի մեջ: Ողջուկը պարունակում է հիմնականում կոնյուգավորված լուծույթային թթուներ, այսինքն ՝ նրանց միացությունները գլիկինով կամ տաուրինով: Խուղակի մեջ թարախաթթուների կոնցենտրացիան մոտ 1% է: Ներառված է լեղու թթուների հիմնական մասը հեպատենտրոն շրջանառություն:Bեղեղի թթուների մի փոքր մասն `օրական մոտ 0,5 գ, արտազատվում է ոսկրերի մեջ: Սա փոխհատուցվում է լյարդում նոր լեղու թթուների սինթեզով, լեղու թթուների ֆոնդը թարմացվում է մոտ 10 օրվա ընթացքում: Խոլեստերինը արտազատվում է նաև հիմնականում աղիքներով: Այն մտնում է աղիքները սննդով և լյարդից `որպես լեղի մաս: Արյան մեջ ներծծված խոլեստերինը պարունակում է մի կտոր, որը ծագում է լեղակիցէնդոգեն խոլեստերինսինթեզվել է լյարդի մեջ), և սննդից ստացված մասը (էկզոգեն խոլեստերին) Հյուսվածքից խոլեստերինի հեռացումը տեղի է ունենում նրա օքսիդացումով լյարդի աղիքային թթուների հետ, որին հաջորդում է դրանց արտազատումը ֆեկցիաներով (օրական մոտավորապես 0,5 գ) և անփոփոխ խոլեստերինի արտազատմամբ (նաև ֆեկցիաներով): Գրենական վիճակում. (Խոլեստերինվերջ + Խոլեստերիննախկին) - (խոլեստերինարտազատում + Լեղաթթուներարտազատում) = 0 Եթե այս հավասարակշռությունը խանգարում է, ապա հյուսվածքներում և արյան մեջ խոլեստերինի կոնցենտրացիան փոխվում է: Արյան խոլեստերինի բարձրացում հիպերխոլեստերեմիա. Սա մեծացնում է աթերոսկլերոզի և լեղապարկի հիվանդության հավանականությունը: LIPID- ի փոխանակման կանոնակարգ Լիպիդային նյութափոխանակությունը կարգավորվում է կենտրոնական նյարդային համակարգի կողմից: Երկարատև բացասական հուզական սթրեսը, կատեխոլամինների արտազատման ավելացումը արյան մեջ կարող է առաջացնել նկատելի քաշի կորուստ: Գործողություն գլյուկագոն լիպոլիտիկ համակարգի վրա նման է կատեչոլամինների գործողությանը: Adrenaline և norepinephrine բարձրացնել հյուսվածքների լիպազայի ակտիվությունը և ճարպային հյուսվածքում լիպոլիզի արագությունը, ինչի արդյունքում արյան պլազմայում ավելանում է ճարպաթթուների պարունակությունը: Ինսուլին ունի ադրենալինի և գլյուկագոնի հակառակ ազդեցությունը լիպոլիզի և ճարպաթթուների մոբիլիզացման վրա: Աճման հորմոն խթանում է լիպոլիզը ՝ դրդելով ացենիլատիկ ցիկլազի սինթեզը: Հիպոֆիզի հիպոֆունկցիան հանգեցնում է մարմնում ճարպի տեղակայմանը (մարսողական գիրություն): Տիրոքսինը, սեռական հորմոններընույնպես ազդում են լիպիդային նյութափոխանակության վրա: Կենդանիների մոտ սեռական խցուկների հեռացումը առաջացնում է ավելորդ ճարպաթափում: LIPID METABOLIC DISORDERS Խոլեստերինը կենդանիների օրգանիզմներին հատուկ ստերոիդ է: Մարդու մարմնում դրա ձևավորման հիմնական տեղը լյարդն է, որտեղ սինթեզվում է խոլեստերինի 50% -ը, 15-20% -ը ձևավորվում է փոքր աղիքի մեջ, մնացածը սինթեզվում է մաշկի մեջ, վերերիկամային ծառի կեղեվ և գոնադներ: Խոլեստերինի ֆոնդի ձևավորման աղբյուրները և դրա ծախսման եղանակները ներկայացված են Նկար 22.1-ում: Նկ. 22.1. Մարմնում խոլեստերինի ձևավորում և բաշխում: Մարդու մարմնի խոլեստերինը (ընդհանուր քանակությունը մոտ 140 գ) պայմանականորեն կարելի է բաժանել երեք լողավազանների. 30 գ), որը արագ փոխանակվում է, բաղկացած է աղիքային պատի խոլեստերինից, արյան պլազմայից, լյարդից և պարենխիմալ այլ օրգաններից, նորացումը տեղի է ունենում 30 օրվա ընթացքում (1 գ / օր), 50 գ), դանդաղորեն փոխանակելով այլ օրգանների և հյուսվածքների խոլեստերինը, 60 գ), ողնաշարի և ուղեղի շատ դանդաղ փոխանակված խոլեստերին, կապի հյուսվածքներ, թարմացման արագությունը հաշվարկվում է տարիների ընթացքում: Խոլեստերինի սինթեզը տեղի է ունենում բջիջների ցիտոզոլում: Սա մարդու մարմնի ամենաերկար նյութափոխանակության ուղիներից մեկն է: Այն ընթանում է 3 փուլով. Առաջինը ավարտվում է մեվալոնաթթվի ձևավորմամբ, երկրորդը `սկվալինի ձևավորմամբ (գծային ածխաջրածին կառուցվածք, որը բաղկացած է 30 ածխածնի ատոմներից): Երրորդ փուլի ընթացքում սկկալինը վերածվում է լանոստերոլի մոլեկուլի, այնուհետև կան 20 հաջորդական ռեակցիա, որոնք լանոստերոլը վերածում են խոլեստերինի: Որոշ հյուսվածքներում խոլեստերինի հիդրօքսի խումբը էստերիֆիկացնում է էստերներ ձևավորելու համար: Արձագանքը կատալիզացվում է AHAT- ի ներբջջային ֆերմենտով (acylCoA: խոլեստերին acyltransferase): The esterification ռեակցիան նույնպես տեղի է ունենում արյան մեջ HDL- ում, որտեղ տեղակայված է ֆերմենտը LHAT (լեցիտին. Խոլեստերինի ացիլtransferase): Խոլեստերինի եթերները այն ձևն են, որով այն տեղափոխվում է արյան միջոցով կամ պահվում է բջիջներում: Արյան մեջ խոլեստերինի մոտ 75% -ը եթերների տեսքով է: Խոլեստերինի սինթեզը կարգավորվում է ՝ ազդելով գործընթացի հիմնական ֆերմենտի գործունեության և քանակի վրա ՝ 3-հիդրօքսի-3-մեթիլգլուտարիլ-CoA ռեդուկտազ (HMG-CoA reductase): Դա ձեռք է բերվում երկու եղանակով. 1. HMG-CoA ռեդուկտազի ֆոսֆորիլացիա / dephosphorylation: Ինսուլինը խթանում է HMG-CoA reductase- ի dephosphorylation- ը ՝ դրանով իսկ այն վերածելով ակտիվ վիճակի: Հետևաբար, կլանման շրջանում խոլեստերինի սինթեզը մեծանում է: Այս ժամանակահատվածում մեծանում է նաև սինթեզի համար ՝ մեկնարկային սուբստրատի առկայությունը: Գլյուկագոնն ունի հակառակ ազդեցություն. A սպիտակուցային kinase- ի միջոցով խթանում է HMG-CoA ռեդուկտազի ֆոսֆորիլացումը ՝ այն վերածելով ոչ ակտիվ վիճակի: Արդյունքում, հետանցքային ժամանակահատվածում և ծոմապահության ժամանակ խոլեստերինի սինթեզը խոչընդոտվում է: 2. HMG-CoA reductase- ի սինթեզի խոչընդոտում:Խոլեստերինը (նյութափոխանակության ուղու վերջնական արտադրանք) նվազեցնում է HMG-CoA reductase գենի արտագրման տոկոսադրույքը ՝ դրանով իսկ խոչընդոտելով սեփական սինթեզը, և լեղաթթուները նույնպես առաջացնում են նմանատիպ ազդեցություն: Արյան խոլեստերինի տեղափոխումը իրականացվում է որպես դեղամիջոցի մի մաս: LP- ները հյուսվածքներում ապահովում են էկզոգեն խոլեստերին, որոշում են նրա հոսքերը օրգանների և մարմնից արտազատման միջև: Էկզոգեն խոլեստերինը հանձնվում է լյարդին ՝ որպես մնացորդային ChM- ի մաս: Այնտեղ, սինթեզված էնդոգեն խոլեստերինի հետ միասին, ստեղծում է ընդհանուր ֆոնդ: Հեպատոցիտներում TAG- ը և խոլեստերինը փաթեթավորված են VLDL- ում, և այս ձևով գաղտնի են արյան մեջ: Արյան մեջ, VLDL- ն LP-lipase- ի գործողության ներքո, որը հիդրոլիզացնում է TAG- ը գլիցերինի և ճարպաթթուների նկատմամբ, նախ փոխարկվում է LSPP, իսկ հետո `LDL, որը պարունակում է մինչև 55% խոլեստերին և դրա եթերներ: LDL- ը խոլեստերինի հիմնական տրանսպորտային ձևն է, որում այն առաքվում է հյուսվածքներ (խոլեստերինի 70% -ը և արյան մեջ նրա էսթերները LDL- ի մի մասն են): Արյունից LDL- ը մտնում է լյարդը (մինչև 75%) և այլ հյուսվածքներ, որոնք ունեն իրենց մակերևույթի վրա LDL ընկալիչները: Եթե բջիջ մտնող խոլեստերինի քանակը գերազանցում է դրա կարիքը, ապա ճնշվում է LDL ընկալիչների սինթեզը, ինչը նվազեցնում է արյան մեջ խոլեստերինի հոսքը: Ընդհակառակը, բջիջում ազատ խոլեստերինի կոնցենտրացիայի նվազումով, ընդհակառակը, ակտիվանում է ընկալիչների սինթեզը: Հորմոնները ներգրավված են LDL ընկալիչների սինթեզի կարգավորմանը. Ինսուլինը, տրիիոդոթիրոնինը և սեռական հորմոնները մեծացնում են ընկալիչների ձևավորումը, իսկ գլյուկոկորտիկոիդները նվազում են: Այսպես կոչված «խոլեստերինի վերադարձի տրանսպորտում», այսինքն. այն ճանապարհը, որն ապահովում է խոլեստերինի վերադարձը լյարդին, հիմնական դեր է խաղում HDL- ն: Դրանք սինթեզվում են լյարդի մեջ `անասելի պրեկուրսորների տեսքով, որոնք գործնականում չեն պարունակում խոլեստերին և TAG: Արյան մեջ HDL- ի նախադրյալները հագեցած են խոլեստերինով ՝ այն ստանալով այլ LP- ներից և բջջային թաղանթներից: Խոլեստերին HDL- ի տեղափոխումը ներառում է դրանց մակերեսին տեղակայված LHAT ֆերմենտ: Այս ֆերմենտը կցում է ճարպաթթվի մնացորդը `ֆոսֆատիդիլխոլինից (լեցիտին) մինչև խոլեստերին: Արդյունքում ձևավորվում է խոլեստերինի էստերի հիդրոֆոբ մոլեկուլ, որը շարժվում է HDL- ի ներսում: Այսպիսով, ոչ հարբած HDL- ը, հարստացված խոլեստերինով, վերածվում է HDL 3-ի - հասուն և ավելի մեծ մասնիկներ: HDL 3 փոխանակել խոլեստերինի էթերներ TAG- ում պարունակվող VLDL և STD- ում `հատուկ սպիտակուցի մասնակցությամբ, որը խոլեստերինի էթերները փոխանցում է լիպոպրոտեինների միջև: Այս դեպքում ՝ HDL 3 վերածվել HDL2- ի, որի չափը մեծանում է TAG կուտակման պատճառով: LLD-lipase- ի ազդեցության տակ VLDL- ը և STDL- ը վերածվում են LDL- ի, որոնք հիմնականում խոլեստերին են մատակարարում լյարդին: Խոլեստերինի մի փոքր մասն առաքվում է HDL2 և HDL լյարդին: Լեղու թթուների սինթեզ: Լյարդում, օրական 5-700 մգ աղիքային թթուներ են սինթեզվում խոլեստերինից: Դրանց ձևավորումը ներառում է հիդրոքսիլազների մասնակցությամբ հիդրոքսիլային խմբերի ներդրման և խոլեստերինի կողմնակի շղթայի մասնակի օքսիդացման ռեակցիաներ (Նկար 22.2). Նկ. 22.2. Լեղու թթուների ձևավորման սխեման: Սինթեզի առաջին ռեակցիան ՝ 7-ա-հիդրօքսիխոլեստերինի ձևավորումը, կարգավորիչ է: Այս ռեակցիան կատալիզացնող ֆերմենտի գործունեությունը խանգարվում է ճանապարհի վերջնական արտադրանքի ՝ աղիների թթուներով: Կարգավորման մեկ այլ մեխանիզմ է ֆերմենտի ֆոսֆորիլացումը / դեֆոսֆորիլացումը (7-ա-հիդրոքսիլազի ֆոսֆորիլացված ձևը ակտիվ է): Կարգավորումը հնարավոր է նաև ֆերմենտի քանակը փոխելով. Խոլեստերինը առաջացնում է 7-a-հիդրոքսիլազ գենի արտագրում, իսկ լեղապարկի թթուները ճնշվում են: Վահանաձև գեղձի հորմոնները առաջացնում են 7-ա-հիդրոքսիլազի սինթեզը և էստրոգենի ճնշումը: Էստրոգենի նման ազդեցությունը լեղաթթուների սինթեզի վրա բացատրում է, թե ինչու է լեղապարկի հիվանդությունը կանանց մոտ 3-4 անգամ ավելի հաճախ առաջանում, քան տղամարդկանց մոտ: Խոլեստերինից ձևավորված խոլիկ և ցենոդեօքսիխոլաթթուները կոչվում են «առաջնային աղու թթուներ»: Այս թթուների մեծ մասը անցնում է կոնյուկտացիա `գլիկինի կամ տաուրինի մոլեկուլների ավելացում լեղաթթվի կարբոքսիլ խմբին: Կնկացումը սկսվում է լեղաթթուների ակտիվ ձևի ձևավորման միջոցով - CoA- ի ածանցյալներ, ապա կցվում են տաուրինը կամ գլիկինը, և արդյունքում ձևավորվում են կոնյուկատների 4 տարբերակներ ՝ տաուրոչոլիկ և տաուրենոդեօքսիխոլիկ, գլիկոքոլիկ և գլիկենոդեօքսիխաթթուներ: Դրանք զգալիորեն ավելի ուժեղ էմուլգատորներ են, քան բնօրինակ աղու թթուները: Գլիկինի հետ կոնյուկները ձևավորվում են 3 անգամ ավելին, քան տաուրինի հետ, քանի որ մարմնում տաուրինի քանակը սահմանափակ է: Աղիքներում մանրէազերծող ֆերմենտների գործողության ներքո գտնվող առաջնային լեղու թթուների կոնյուկտները փոքրացվում են երկրորդական լեղու թթուների: Դեօքսիխոլաթթուն, որը կազմված է խոլիկից և լիտոքոլիկից, որը ձևավորվել է դեօքսիխոլից, ավելի քիչ լուծելի է և ավելի դանդաղ կլանում է աղիքները: Լեղու թթուների մոտ 95% -ը, որոնք աղիք են մտնում, վերադառնում են լյարդին պորտալարի երակային միջոցով, այնուհետև կրկին գաղտնազերծվում են լեղապարկի մեջ և կրկին օգտագործում են ճարպերի էմուլգացման մեջ: Լեղաթթուների այս ուղին կոչվում է enterohepatic շրջանառություն: Fարպերով երկրորդային լեղու թթուները հիմնականում հանվում են: Լեղապարկի հիվանդությունը (խոլելիտիասիս) պաթոլոգիական գործընթաց է, որի ընթացքում քարերը ձևավորվում են լեղապարկի մեջ, որի հիմքը խոլեստերինն է: Խոլեստերին խցուկի մեջ արտազատումը պետք է ուղեկցվի լեղաթթուների և ֆոսֆոլիպիդների համաչափ ազատմամբ, որոնք հիփրոֆոբ խոլեստերինի մոլեկուլները պահպանում են միկլարային վիճակում: Bրային թթուների և խոլեստերինի խոլեստերինի հարաբերակցության փոփոխության պատճառ հանդիսացող պատճառներն են ՝ խոլեստերինով հարուստ սնունդ, բարձր կալորիականությամբ սնուցում, լեղու աղիքի լեղու լճացում, աղտոտված enterohepatic շրջանառություն, լեղաթթուների թուլացման սինթեզ, լեղապարկի վարակ: Խոլելիտիասի հիվանդությամբ հիվանդների մեծ մասում խոլեստերինի սինթեզն աճում է, և դրանից ստացվում է լեղաթթուների սինթեզը, ինչը հանգեցնում է աղիների մեջ արտանետվող խոլեստերինի և լեղու թթուների քանակի անհամաչափությանը: Արդյունքում, խոլեստերինը սկսում է նստել լեղապարկի մեջ, ձևավորելով մածուցիկ նստվածք, որն աստիճանաբար կարծրացնում է: Երբեմն այն ներմուծվում է բիլիրուբինով, սպիտակուցներով և կալցիումի աղերով: Քարերը կարող են բաղկացած լինել միայն խոլեստերինից (խոլեստերինի քարեր) կամ խոլեստերինի, բիլիրուբինի, սպիտակուցների և կալցիումի խառնուրդ: Խոլեստերինի քարերը սովորաբար սպիտակ են, իսկ խառը քարերը տարբեր ստվերներով շագանակագույն են: Քարի ձևավորման սկզբնական փուլում ցենոդոքսիկոլաթթուն կարող է օգտագործվել որպես դեղամիջոց: Երբ լեղապարկի մեջ լինելով, այն աստիճանաբար լուծարում է խոլեստերինի քարերը, բայց սա դանդաղ գործընթաց է, որը տևում է մի քանի ամիս: Աթերոսկլերոզը պաթոլոգիա է, որը բնութագրվում է անոթային պատի ներքին մակերեսի վրա աթերոգենային սալերի տեսքով: Նման պաթոլոգիայի զարգացման հիմնական պատճառներից մեկն այն է, որ սննդից խոլեստերին խլելու, դրա սինթեզի և մարմնից արտազատման միջև հավասարակշռության խախտում է: Աթերոսկլերոզով հիվանդները բարձրացրել են LDL և VLDL կոնցենտրացիաները: HDL կոնցենտրացիայի և աթերոսկլերոզի առաջացման հավանականության միջև կա փոխադարձ կապ: Սա համահունչ է LDL- ի գործողության հայեցակարգին, որպես հյուսվածքներում խոլեստերինի կրողներ, իսկ հյուսվածքներից `HDL: Աթերոսկլերոզի զարգացման հիմնական մետաբոլիկ «նախադրյալը» հիպերխոլեստերեմիան է: (արյան մեջ բարձր խոլեստերին): Հիպերխոլեստերինը զարգանում է. 1. խոլեստերինի, ածխաջրերի և ճարպերի ավելցուկի պատճառով. 2. գենետիկ նախատրամադրվածություն, որը բաղկացած է ժառանգական արատներից LDL կամ apoB-100 ընկալիչների կառուցվածքում, ինչպես նաև apoB-100 սինթեզի կամ սեկրեցիայի աճով (ընտանեկան համակցված հիպերլիպիդեմիայի դեպքում, որի դեպքում արյան կոնցենտրացիաները և խոլեստերինը և TAG- ն աճում են): Աթերոսկլերոզի զարգացման մեխանիզմներում կարևոր դեր է խաղում դեղամիջոցի ձևափոխմամբ: LDL- ում լիպիդների և սպիտակուցների բնականոն կառուցվածքի փոփոխությունները դրանք օտար են դարձնում օրգանիզմի համար, ուստի ավելի մատչելի են ֆագոցիտների կողմից գրավման համար: Թմրամիջոցների ձևափոխումը կարող է առաջանալ մի քանի մեխանիզմներով. 1. սպիտակուցների գլիկոզիլացում, որը տեղի է ունենում արյան մեջ գլյուկոզի համակենտրոնացման բարձրացման դեպքում, 2. պերօքսիդի ձևափոխում ՝ հանգեցնելով լիպոպրոտեինների լիպիդների փոփոխության և apoB-100 կառուցվածքի, 3. LP- հակամարմինների աուտոիմունային բարդույթների ձևավորումը (փոփոխված դեղամիջոցները կարող են առաջացնել աուտանտիտների ձևավորում): Փոփոխված LDL- ն ներծծվում է մակրոֆագների միջոցով: Այս գործընթացը չի կարգավորվում կլանված խոլեստերինի քանակով, քանի որ այն հատուկ ռեցեպտորների միջոցով բջիջներ մուտք գործելու դեպքում, հետևաբար մակրոֆագները ծանրաբեռնված են խոլեստերինով և վերածվում են «փրփրացող բջիջների», որոնք ներթափանցում են subendothelial տարածություն: Սա հանգեցնում է արյան անոթների պատի մեջ լիպիդային բծերի կամ շերտերի ձևավորմանը: Այս փուլում անոթային էնդոթելիումը կարող է պահպանել իր կառուցվածքը: Փրփուրի բջիջների քանակի աճով էնդոթելիի վնաս է առաջանում: Վնասը նպաստում է թրոմբոցիտների ակտիվացմանը: Արդյունքում, նրանք գաղտնազերծում են թրոմբոքսանը, որը խթանում է թրոմբոցիտների ագրեգացումը, ինչպես նաև սկսում են արտադրել թրոմբոցիտների ստացման աճի գործոն, ինչը խթանում է հարթ մկանների բջիջների բազմացումը: Վերջիններս մեդալիայից տեղափոխվում են զարկերակային պատի ներքին շերտ ՝ դրանով իսկ նպաստելով սալիկի աճին: Բացի այդ, մանրաթելային հյուսվածքով սալիկապատված թիթեղները, մանրաթելային թաղանթի տակ գտնվող բջիջները նեկրոտիկ են, իսկ խոլեստերինը պահվում է միջբջջային տարածքում: Մշակման վերջին փուլերում սալը ներկված է կալցիումի աղերով և դառնում է շատ խիտ: Սալիկի տարածքում արյան խցանումները հաճախ ձևավորվում են ՝ խցանելով նավի անոթները, ինչը հանգեցնում է համապատասխան հյուսվածքների տեղանքում սուր շրջանառության խանգարման և սրտի կաթվածի զարգացման: Խոլեստերինի սինթեզի կարգավորումը `դրա հիմնական ֆերմենտը (HMG-CoA reductase) իրականացվում է տարբեր եղանակներով: HMG ռեդուկտազի ֆոսֆորիլացիա / dephosphorylation: Ինսուլին / գլյուկագոն հարաբերակցության բարձրացումով, այս ֆերմենտը ֆոսֆորիլացնում է և անցնում ակտիվ վիճակի: Ինսուլինի գործողությունը իրականացվում է 2 ֆերմենտների միջոցով: HMG-CoA reductase kinase phosphatase, որը kinase- ը վերածում է անգործուն dephosphorylated պետության: Phosphotase HMG-CoA reductase- ը այն վերածելով dephosphorylated ակտիվ պետության: Այս ռեակցիաների արդյունքը HMG-CoA ռեդուկտազի դեֆոսֆորիլացված ակտիվ ձևի ձևավորումն է: Հետևաբար, կլանման ժամանակահատվածում խոլեստերինը մեծանում է: Այս ժամանակահատվածում ավելանում է խոլեստերինի `ացետիլ - CoA- ի սինթեզի համար նախնական սուբստրատի առկայությունը (ածխաջրեր և ճարպեր պարունակող սնունդ ուտելու արդյունքում, քանի որ CoA- ի ացետիլը ձևավորվում է գլյուկոզի և ճարպաթթուների տրոհման ժամանակ): Հետցնցումային վիճակում գլյուկագոնը A պրոտեինոգենազի միջոցով խթանում է HMG - CoA- ի ֆոսֆորիլացումը `ռեդուկտազը, այն վերածելով ոչ ակտիվ վիճակի: Այս գործողությունը ուժեղանում է նրանով, որ միևնույն ժամանակ գլյուկագոնը խթանում է HMG-CoA reductase ֆոսֆոտազի ֆոսֆորիլացումը և անգործությունը, դրանով իսկ HMG-CoA- ի ռեդուկտազը պահելով ֆոսֆորիլացված ոչ ակտիվ վիճակում: Արդյունքում, հետբաբսորման ժամանակահատվածում և ծոմապահության ժամանակ խոլեստերինի սինթեզը արգելվում է էնդոգեն սինթեզով: Եթե սննդի մեջ խոլեստերին պարունակությունը բերվել է 2% -ի, ապա էնդոգեն խոլեստերինի սինթեզը կտրուկ նվազել է: Բայց խոլեստերինի սինթեզի ամբողջական դադարեցումը տեղի չի ունենում: Խոլեստերինի բիոսինթեզի արգելակման աստիճանը սննդից եկող խոլեստերինի ազդեցության տակ տատանվում է անձից այլ անձի: Սա ցույց է տալիս խոլեստերինի ձևավորման գործընթացների անհատականությունը: Նվազեցնելով խոլեստերինի սինթեզի ինտենսիվությունը, հնարավոր է նվազեցնել նրա կոնցենտրացիան արյան մեջ: Եթե մի կողմից մարմնում խոլեստերին ընդունողկազմի և մի կողմից մարմնում դրա սինթեզի միջև հավասարակշռությունը խախտվում է, մյուս կողմից `լեղաթթուների և խոլեստերինի արտանետումների միջև, խախտվում է, հյուսվածքներում խոլեստերինի կոնցենտրացիան փոխվում է: Ամենալուրջ հետևանքները կապված են արյան խոլեստերինի համակենտրոնացման բարձրացման հետ (հիպերխոլեստերեմիա), մինչդեռ մեծանում է աթերոսկլերոզի և խոլելիտիասի զարգացման հավանականությունը: Ընտանեկան հիպերխոլեստերեմիա (HCS) - այս ձևը շատ ավելի տարածված է `200 մարդու մոտ 1 հիվանդ: HCS- ում ժառանգված թերությունը բջիջների կողմից LDL- ի կլանման խախտում է, և, հետևաբար, LDL կատաբոլիզմի մակարդակի նվազում: Արդյունքում, արյան մեջ LDL- ի կոնցենտրացիան բարձրանում է, ինչպես նաև խոլեստերինը, քանի որ LDL- ում դրա մեծ մասը կա: Հետևաբար, HCS- ով բնորոշ է հյուսվածքներում խոլեստերինի պահպանումը, մասնավորապես մաշկում (քսանտոմա), զարկերակների պատերին: HMG-CoA ռեդուկտազի սինթեզի խոչընդոտում Խոլեստերինի նյութափոխանակության ուղու վերջնական արտադրանքը: Այն նվազեցնում է HMG-CoA reductase գենի արտագրման տեմպը ՝ դրանով իսկ խանգարելով սեփական սինթեզին: Լյարդը ակտիվորեն սինթեզում է լեղու թթուները խոլեստերինից, և, հետևաբար, լեղաթթուները խանգարում են HMG-CoA reductase գենի գործունեությունը: Քանի որ HMG-CoA ռեդուկտազը գոյություն ունի մոտ 3-ի սինթեզից հետո, այս խոլեստերինի ֆերմենտի սինթեզի արգելակումը արդյունավետ կարգավորում է: |