Խոլեստերինի բիոսինթեզ և դրա կենսաքիմիա `շաքարախտ
Անկասկած, խոլեստերինը հանրության շրջանում ամենատարածված լիպիդն է, այն տխրահռչակ է արյան բարձր խոլեստերինի և մարդու սրտանոթային հիվանդությունների հաճախականության միջև բարձր փոխկապվածության պատճառով: Ավելի քիչ ուշադրություն է դարձվել խոլեստերինի վճռորոշ դերին `որպես բջջային մեմբրանների բաղադրիչ և որպես ստերոիդ հորմոնների և լեղաթթուների նախադրյալ: Խոլեստերինը անհրաժեշտ է շատ կենդանիների, այդ թվում նաև մարդու համար, բայց կաթնասունի սննդի մեջ դրա ներկայությունը կամընտիր է `մարմնի բջիջները իրենք կարող են սինթեզել այն պարզ նախորդներից:
Այս 27-ածխածնի միացության կառուցվածքը ենթադրում է բարդ ուղի իր բիոսինթեզի համար, բայց դրա բոլոր ածխածնի ատոմները տրամադրվում են մեկ պրեկուրսորի կողմից `ացետատով: Isoprene բլոկներ - ամենակարևոր միջնորդները ՝ ացետատից մինչև խոլեստերին, դրանք բնական բնական լիպիդների նախածերներն են, և այն մեխանիզմները, որոնց միջոցով իզոպրենային բլոկները պոլիմերացված են, նման են բոլոր նյութափոխանակության ուղիներով:
Մենք սկսում ենք քննարկել ացետատից խոլեստերինի բիոսինթեզի ուղու հիմնական փուլերը, այնուհետև քննարկում ենք խոլեստերինի տեղափոխումը արյան մեջ, բջիջների կողմից դրա կլանումը, խոլեստերինի սինթեզի նորմալ կարգավորումը և կարգավորվածությունը թուլացած կլանման կամ տեղափոխման դեպքում: Այնուհետև մենք նայում ենք այլ նյութեր, որոնք գալիս են խոլեստերինից, ինչպիսիք են լեղաթթուները և ստերոիդ հորմոնները: Ի վերջո, բազում միացությունների ձևավորման համար կենսասինթետիկ ուղիների նկարագրությունը `իզոպրենային բլոկների ածանցյալներ, որոնցում կան ընդհանուր խոլեստերինի սինթեզի հետ կապված վաղ վաղ փուլեր, կներկայացնի բիոսինթեզում isoprenoid խտացման արտառոց բազմակողմանիությունը:
Խոլեստերինը արտադրվում է ացետիլ-CoA- ից չորս փուլով
Խոլեստերինը, ինչպես երկար շղթայի ճարպաթթուները, պատրաստված է ացետիլ-CoA- ից, բայց հավաքման օրինակը բոլորովին այլ է: Առաջին փորձարկումներում, 14 C- ով պիտակավորված ացետատը, կամ մեթիլային կամ կարբոքսիլ ածխածնի ատոմում, ավելացվել է կենդանիների կեր: Հիմք ընդունելով պիտակի բաշխումը կենդանիների երկու խմբերից մեկուսացված խոլեստերինում (Նկար 21-32), նկարագրվել են խոլեստերինի բիոսինթեզի ֆերմենտային փուլերը:
Նկ. 21-32: Խոլեստերինի ածխածնի ատոմների աղբյուր: Հայտնաբերվել է մեթիլ ածխածնի (սև) կամ կարբոքիլային ածխածնի (կարմիր) պիտակավորված ռադիոակտիվ ացետատ օգտագործող փորձերի ժամանակ: Խտացրած կառուցվածքում օղակները նշվում են A-D տառերով:

Սինթեզը տեղի է ունենում չորս փուլով, ինչպես ցույց է տրված Նկ. 21-33: ստերոիդ կորիզի չորս օղակներ, որոնց հաջորդում են մի շարք փոփոխություններ (օքսիդացում, մեթիլ խմբերի հեռացում կամ միգրացիա) խոլեստերինի ձևավորմամբ:
Նկ. 21-33: Խոլեստերինի բիոսինթեզի ընդհանրացված պատկերը: Տեքստում քննարկվում են սինթեզի չորս փուլերը: Սկալալենում գտնվող իզոպրենային բլոկները նշվում են կարմիր գծապատկերային գծերով:

Բեմ (1): Մեզալոնատի սինթեզ ացետատից: Խոլեստերինի բիոսինթեզի առաջին փուլը հանգեցնում է միջանկյալ արտադրանքի ձևավորմանը մևալոնացնել (Նկար 21-34): Երկու ացետիլ CoA մոլեկուլը խտացնում է ացետացետիլ CoA- ին տալը, որը երրորդ ացետիլ CoA մոլեկուլով կոնդենսացնում է վեց ածխածնի միացություն կազմելու համար β-հիդրօքսի-β-մեթիլգլուտարիլ-CoA (HM G -CoA): Այս երկու առաջին ռեակցիաները կատալիզացված են տիոլազ համապատասխանաբար և NM G -CoA սինթազ: Ytիտոզոլիկ NM G-CoA սինթազ Այս նյութափոխանակության ճանապարհը տարբերվում է միիտոկոնդրիալ իզոեզիմից, որը կատալիզացնում է NM G-CoA- ի սինթեզը ketone մարմինների ձևավորման ժամանակ (տես Նկար 17-18):
Նկ. 21-34: Մեթվոլոնատի ձևավորումը ացետիլ-CoA- ից: Ացետիլ-CoA- ից C-1 և C-2 mevalonate- ի աղբյուրը կարևորվում է վարդագույն գույնով:

Երրորդ արձագանքը սահմանափակում է ամբողջ գործընթացի արագությունը: Դրա մեջ NM G -CoA- ն իջեցվում է mevalonate- ի, որի համար NA D PH մոլեկուլներից յուրաքանչյուրը տրամադրում է երկու էլեկտրոն: HMG-CoA reductase - սահուն ER- ի ինտեգրալ թաղանթային սպիտակուցը, այն ծառայում է, ինչպես հետագայում կտեսնենք, որպես խոլեստերինի ձևավորման նյութափոխանակային ուղու կարգավորման հիմնական կետ:
Բեմ (2): Մեվալոնատի վերածումը երկու ակտիվացված իզոպրենի: Խոլեստերինի սինթեզի հաջորդ փուլում երեք ֆոսֆատ խմբերը ATP մոլեկուլներից տեղափոխվում են մեվալոնատ (նկ. 21-35): Միջին 3-ֆոսֆո-5-պիրոֆոսֆոմեվոլոնատում հիդրոքսիլային խմբին միացված հիդրոքսիլային խմբին կապված ֆոսֆատը լավ հեռացող խումբ է, հաջորդ քայլում և այս ֆոսֆատներից, և հարակից կարբոքսիլային խմբից մեկնում են երկքաղաքային կապ:իզոպանտենիլ պիրոֆոսֆատ: Սա ակտիվացված երկու իզոպրեններից առաջինն է `խոլեստերինի սինթեզի հիմնական մասնակիցները: Δ 3-իզոպանտենիլպիրոֆոսֆատի իզոմերացումը տալիս է երկրորդ ակտիվացված իզոպրեն dimethylallyl պիրոֆոսֆատ: Բույսերի բջիջների ցիտոպլազմում իզոպենտենիլ պիրոֆոսֆատի սինթեզը տեղի է ունենում այստեղ նկարագրված ուղու համաձայն: Այնուամենայնիվ, բույսերի քլորոպլաստները և շատ մանրէներ օգտագործում են մևալոնատից անկախ ուղի: Այս այլընտրանքային ուղին կենդանիների մոտ չի հայտնաբերվում, ուստի այն գրավիչ է նոր հակաբիոտիկների ստեղծման ժամանակ:
Նկ. 21-35: Մեվալոնատի վերափոխումը ակտիվացված իզոպրենի բլոկների: Վեց ակտիվացված միավորները միավորվում են սկալալեն կազմելու համար (տես Նկար 21-36): 3-ֆոսֆո-5-պիրոֆոսֆոմեվոլոնատի հեռացող խմբերը կարևորվում են վարդագույն գույնով: Քառակուսի փակագծերում հիպոթետիկ միջնորդ է:

Բեմ (3): Վեց ակտիվացված իզոպրեն միավորների խտացում `սկկալեն կազմելու համար: Իզոպենթենիլ պիրոֆոսֆատը և dimethylallyl պիրոֆոսֆատը այժմ անցնում են գլխի-պոչի խտացում, որում շարժվում է մեկ պիրոֆոսֆատ խումբ և 10-ածխածնի շղթա ձևավորվում է - geranyl պիրոֆոսֆատ (Նկար 21-36): (Պիրոֆոսֆատը կցվում է գլխին): Գերանիլ պիրոֆոսֆատը անցնում է հետևից գլուխ-պոչի խտացում իզոպանտենիլ պիրոֆոսֆատով և 15-ածխածնի միջանկյալ ձևերով ֆարնեսիլ պիրոֆոսֆատ: Վերջապես, ֆարնեսիլ պիրոֆոսֆատի երկու մոլեկուլը համատեղում է «գլուխը դեպի գլուխ», ֆոսֆատ խմբերը հանվում են. սկվալեն:
Նկ. 21-36: Squalene- ի ձեւավորում: Սքվալենի կառուցվածքը, որը պարունակում է 30 ածխածնի ատոմ, տեղի է ունենում իզոպրենի (հինգ-ածխածնի) բլոկների ակտիվացված հաջորդական խտացման ընթացքում:

Այս միջնորդների համար ընդհանուր անունները գալիս են այն աղբյուրների անուններից, որոնցից նրանք առաջին անգամ մեկուսացվել են: Գերանիոլը, վարդի յուղի բաղադրիչ, խորդենի բույր ունի, և ֆառնեսոլը, որը հայտնաբերված է ակացիա ֆարնեսայի գույներով, ունի հովտի բույր: Բույսերի շատ բնական հոտեր պատկանում են իզոպրենային բլոկներից կառուցված միացություններին: Սկալալենը, որն առաջին անգամ մեկուսացված է շնաձկների լյարդից (Squalus տեսակներ), բաղկացած է 30 ածխածնի ատոմից. Հիմնական շղթայում 24 ատոմ և մետաղական փոխարինողներից վեց ատոմ:
Բեմ (4): Սկալալենի վերափոխումը ստերոիդ կորիզի չորս օղակների. Նկ. 21-37-ին ակնհայտորեն երևում է, որ սկվալենի շղթայի կառուցվածքը, իսկ ստերոլները ՝ ցիկլային: Բոլոր ստերոլները ունեն չորս խտացրած օղակ, որոնք կազմում են ստերոիդային կորիզը, և բոլորն ալ C- 3 ատոմում հիդրոքսիլ խմբով ալկոհոլներ են, հետևաբար անգլիական ստերոլ անունն է: Գործողության ներքո squalene monooxygenase O- ից մեկ թթվածնի ատոմ ավելացվում է սպորտային շղթայի ավարտին 2 և ձևավորվում է էպօքսիդ: Այս ֆերմենտը ևս մեկ խառը գործառույթով օքսիդազ է (ավելացրեք 21-1), NADPH- ն իջեցնում է թթվածնի մեկ այլ ատոմ O- ից 2 դեպի Հ2 Օ. Ապրանքի կրկնակի կապեր squalene-2,3-epoxide կազմակերպված այնպես, որ զգալիորեն հետևողական արձագանքը կարող է սկալլեն էպօքսիդի շղթան վերածել ցիկլային կառուցվածքի: Կենդանիների բջիջներում այս ցիկլիզացումը հանգեցնում է դրանց ձևավորմանը լանոստերոլ որը պարունակում է չորս օղակներ, որոնք բնորոշ են ստերոիդային կորիզին: Արդյունքում, լանոստերոլը վերածվում է խոլեստերին ՝ մոտավորապես 20 ռեակցիաների շարքի միջոցով, որը ներառում է որոշ մետաղական խմբերի միգրացիա և այլոց հեռացում: Կենսազինթեզի այս զարմանալի ուղու նկարագրությունը, որը հայտնիներից ամենադժվարներից մեկն էր, կազմել են Քոնրադ Բլոչը, Թեոդոր Լինենը, Johnոն Քորնֆորը և Georgeորջ Պոպիակը 1950-ականների վերջին:
Նկ. 21-37: Օղակների փակումը գծային սկալենը վերածում է խտացրած ստերոիդային միջուկի: Առաջին փուլը կատալիզացվում է խառը գործառույթով օքսիդազազով (մոնոօքսիգենազ), որի տրոհման հիմքն է N AD PH: Ապրանքը էպօքսիդ է, որը հաջորդ փուլում ցիկլիզացվում է ՝ ստերոիդային միջուկ ձևավորելու համար: Կենդանիների բջիջներում այս ռեակցիաների վերջնական արտադրանքը խոլեստերինն է. Այլ օրգանիզմներում ձևավորվում են նրանից ստերոլներ:


Խոլեստերինը ստերոլ է, որը բնորոշ է կենդանական բջիջներին, բույսերը, սնկերը և պրոտիստները արտադրում են այլ շատ նման ստերոլներ:
Նրանք օգտագործում են նույն սինթեզման ուղին դեպի սկվալեն-2,3-էօքսիդ, բայց այնուհետև ուղիները մի փոքր շեղվում են, և ձևավորվում են այլ ստերոլներ, ինչպիսիք են սիգմաստերոլը շատ բույսերում և սնկերի էրգոստերոլը (սնկեր)
Օրինակ 21-1 էներգիայի ծախսերը Squalene սինթեզի համար
Որո՞նք են էներգիայի ծախսերը (արտահայտված որպես ATP մոլեկուլ) մեկ սկկալենի մոլեկուլի սինթեզի համար:
Լուծում: Acetyl-CoA- ից squalene- ի սինթեզում ATP- ն ծախսվում է միայն այն փուլում, երբ mevalonate- ը վերածվում է ակտիվացված իզոպրենի սկալենային նախածանի: Սկալալինի մոլեկուլ կառուցելու համար անհրաժեշտ է վեց ակտիվացված իզոպրեն մոլեկուլ, և յուրաքանչյուր ակտիվացված մոլեկուլ արտադրելու համար պահանջվում է երեք ATP մոլեկուլ: Ընդհանուր առմամբ, 18 ATP մոլեկուլ է ծախսվում մեկ սկկալենի մոլեկուլի սինթեզի վրա:
Խոլեստերինի միացություններ մարմնում
Ողնաշարավորներում լյարդում մեծ քանակությամբ խոլեստերին են սինթեզվում: Այնտեղ սինթեզված խոլեստերինի մի մասը ներառված է հեպատոցիտների մեմբրանների մեջ, բայց այն հիմնականում արտահանվում է նրա երեք ձևերից մեկում ՝ աղիքային (լեղու) խոլեստերին, լեղու թթուներ կամ խոլեստերինի էթերեր: Լեղաթթուներ և դրանց աղերը խոլեստերինի հիդրոֆիլային ածանցյալներ են, որոնք սինթեզվում են լյարդի մեջ և նպաստում են լիպիդների մարսմանը (տես Նկար 17-1): Խոլեստերինի էստերներ ձևավորվել է լյարդում `գործողության միջոցով acyl-CoA- խոլեստերին-acyltransferase (ACAT): Այս ֆերմենտը կատալիզացնում է ճարպաթթվի մնացորդի փոխանցումը կոենզիմ A- ից խոլեստերինի հիդրոքսիլ խմբին (Նկար 21-38) ՝ խոլեստերինը վերածելով ավելի հիդրոֆոբ ձևի: Խոլեստերինի եթերները գաղտնի լիպոպրոտեինային մասնիկների մեջ տեղափոխվում են այլ հյուսվածքներ ՝ օգտագործելով խոլեստերին կամ պահվում են լյարդի մեջ:
Նկ. 21-38: Խոլեստերինի էսթերի սինթեզ: Եթերացումը խոլեստերինը դարձնում է ավելի հիդրոֆոբիկ ձև ՝ պահեստավորման և տեղափոխման համար:

Խոլեստերինը անհրաժեշտ է աճող կենդանիների օրգանիզմի բոլոր հյուսվածքների համար մեմբրանների սինթեզի համար, իսկ որոշ օրգաններ (օրինակ ՝ վերերիկամային խցուկներ և սեռական խցուկներ) խոլեստերինը օգտագործում են որպես ստերոիդ հորմոնների նախածանց (այս մասին կքննարկվի ստորև): Խոլեստերինը նաև հանդիսանում է վիտամին D- ի նախադրյալ (տես Նկար 10-20, v. 1):
Խոլեստերինը և այլ լիպիդները կրում են պլազմային լիպոպրոտեիններ
Խոլեստերինի և խոլեստերինի էսթերները, ինչպես տրիասիլգլիցերինները և ֆոսֆոլիպիդները, գործնականում ջրի մեջ անլուծելի են, այնուամենայնիվ, նրանք պետք է տեղափոխվեն այն հյուսվածքից, որի մեջ դրանք սինթեզվել են այն հյուսվածքների մեջ, որտեղ դրանք կպահպանվեն կամ սպառվեն: Դրանք տեղափոխվում են արյան հոսքով `ձևով արյան պլազմային լիպոպրոտեիններ - հատուկ կրիչ սպիտակուցների մակրոմոլեկուլային համալիրներ (ապոլիպոպրոտեիններ) ֆոսֆոլիպիդներով, խոլեստերինով, խոլեստերինի էթերներով և տրիքիիլգլիցերիններով, որոնք առկա են այս համալիրներում տարբեր համակցություններով:
Ապոլիպոպրոտեինները («apo» -ը վերաբերում է լիպիդներից ազատ սպիտակուցին ինքնին) համատեղվում են լիպիդների հետ `կազմելով լիպոպրոտեինային մասնիկների մի քանի ֆրակցիաներ` կենտրոնում գտնվող հիդրոֆոբիկ լիպիդներով գնդաձև բարդույթներ և մակերևույթին գտնվող հիդրոֆիլային ամինաթթուների շղթաներ (Նկար 21-39, ա): Լիպիդների և սպիտակուցների տարբեր համակցություններով ձևավորվում են տարբեր խտությունների մասնիկներ `քիլոմիկրոններից մինչև բարձր խտության լիպոպրոտեիններ: Այս մասնիկները կարելի է առանձնացնել ուլտրաձայնազերծմամբ (Աղյուսակ 21-1) և տեսողականորեն դիտարկել էլեկտրոնային մանրադիտակի միջոցով (Նկար 21-39, բ): Լիպոպրոտեինների յուրաքանչյուր բաժին կատարում է հատուկ գործառույթ, որը որոշվում է սինթեզի, լիպիդային կազմի և ապոլիպոպրոտեինների պարունակությամբ: Առնվազն 10 տարբեր ապոլիպոպրոտեիններ են հայտնաբերվել մարդու արյան պլազմայում (աղյուսակ 21-2), որոնք տարբերվում են չափերով, հատուկ հակամարմիններով արձագանքներով և լիպոպրոտեինների տարբեր դասերում բնութագրական բաշխմամբ: Այս սպիտակուցային բաղադրիչները գործում են որպես ազդանշանային նյութեր, որոնք ուղղորդում են լիպոպրոտեինները դեպի հատուկ հյուսվածքներ կամ ակտիվացնում են ֆերմենտները, որոնք գործում են լիպոպրոտեինների վրա:
Աղյուսակ 21-1: Մարդու պլազմային լիպոպրոտեիններ
Բաղադրությունը (զանգվածային բաժին,%)
r = 513,000): LDL- ի մի մասն իր մեջ պարունակում է խոլեստերինի էսթերների մոտավորապես 1,500 մոլեկուլ, որի առանցքի շուրջ կա խոլեստերինի 500 մոլեկուլ, 800 մոլեկուլ ֆոսֆոլիպիդներ և apoB-100- ի մեկ մոլեկուլ: բ - լիպոպրոտեինների չորս դաս, տեսանելի էլեկտրոնային մանրադիտակով (բացասականության դրսևորումից հետո): Ժամացույցի սլաքի ուղղությամբ, սկսած վերևի ձախ գծից ՝ քիլոմիկրոններ ՝ 50-ից 200 նմ տրամագծով, PL O NP ՝ 28-ից 70 նմ, HDL ՝ 8-ից 11 նմ, իսկ LDL ՝ 20-ից 55 նմ: Լիպոպրոտեինների հատկությունները տրված են աղյուսակում: 21-2:

Chylomicrons, հիշատակված է Sec. 17, սննդի տրիխիլգլիցերինները տեղափոխել աղիքից դեպի այլ հյուսվածքներ: Սրանք ամենամեծ լիպոպրոտեիններն են, նրանք ունեն ամենացածր խտությունը և տրիասկիլգլիցերինների ամենաբարձր հարաբերական պարունակությունը (տե՛ս Նկ. 17-2): Chylomicrons- ը սինթեզվում է էպիթելի բջիջների ER- ում, որոնք պառկեցնում են փոքր աղիքները, այնուհետև շարժվում են ավշային համակարգի միջով և մուտք են գործում արյան հոսք ձախ ենթակլավային երակային միջոցով: Chylomicron apolipoproteins- ը պարունակում է apoB-48 (լիպոպրոտեինների այս դասի համար եզակի), apoE և apoC-II (աղյուսակ 21-2): AroC-II- ն ակտիվացնում է լիպոպրոտեինային լիպազը ճարպային հյուսվածքի, սրտի, կմախքի մկանների և կաթնաթթվային կաթնային գեղձերի մազանոթներում ՝ ապահովելով ազատ ճարպաթթուների հոսքը այդ հյուսվածքներում: Այսպիսով, քիլոմիկրոնները սննդի ճարպաթթուները տեղափոխում են հյուսվածքներ, որտեղ դրանք սպառվելու կամ պահվելու են որպես վառելիք (Նկար 21-40): Chylomicron- ի մնացորդները (հիմնականում ազատված են տրիասիլգլիցերիներից, բայց դեռ պարունակում են խոլեստերին, apoE և apoB-48), որոնք արյան միջոցով տեղափոխվում են լյարդ: Լյարդում ընկալիչները կապում են քիլոմիկրոնի մնացորդներում պարունակվող apoE- ին և միջնորդում են դրանց կլանումը էնդոցիտոզով: Հեպատոցիտներում այս մնացորդները ազատում են իրենց պարունակած խոլեստերինը և ոչնչացվում են լիզոսոմներում:
Աղյուսակ 21-2: Մարդու պլազմային լիպոպրոտեինային ապոլիպոպրոտեիններ
Գործառույթ (եթե հայտնի է)
Ակտիվացնում է L CAT- ը, համագործակցում է ABC փոխադրողի հետ
Խանգարում է L CAT- ին
Ակտիվացնում է L CAT- ը, խոլեստերինի տեղափոխումը / մաքրումը
Կապում է LDL ընկալիչին
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Սկսում է VLDL և քիլոմիկրոնի մնացորդների մաքրում
Երբ սնունդն ավելի շատ ճարպաթթուներ է պարունակում, քան ներկայումս կարող է օգտագործվել որպես վառելիք, դրանք վերածվում են լյարդի տրիասիլգլիցերինների, որոնք կազմում են մասնաբաժին հատուկ ապոլիպոպրոտեիններով: շատ ցածր խտության լիպոպրոտեիններ (VLDL): Լյարդի ավելցուկային ածխաջրերը նույնպես կարող են վերափոխվել տրիասիլգլիցերինների և արտահանվել որպես VLDL (Նկար 21-40, ա):Բացի triacylglycerols- ից, VLDL կոտորակը պարունակում է որոշակի քանակությամբ խոլեստերին և խոլեստերինի էթերներ, ինչպես նաև apoB-100, apoC-1, apoC-II, apoC III և apoE (աղյուսակ 21-2): Այս լիպոպրոտեինները արյան միջոցով տեղափոխվում են նաև լյարդից դեպի մկաններ և ճարպային հյուսվածքներ, որտեղ, լիպոպրոտեինների լիպազը ակտիվացնելուց հետո apo-C II- ով, ազատ ճարպաթթուները ազատվում են VLDL ֆրակցիայի տրիասիլգլիցերիներից: Ադիպոցիտները գրավում են անվճար ճարպաթթուները, դրանք կրկին վերածվում են տրիխիլգլիցերինների, որոնք պահվում են այդ բջիջներում լիպիդային ներառումների (կաթիլների) տեսքով, միոկիտները, ընդհակառակը, անմիջապես օքսիդացնում են ճարպաթթուները ՝ էներգիա ստեղծելու համար: VLDL- ի մնացորդների մեծ մասը շրջանառությունից վերացվում են հեպատոցիտներով: Դրանց կլանումը, որը նման է կիլոմիկրոնների կլանմանը, միջնորդվում է ընկալիչների կողմից և կախված է VLDL մնացորդներում apoE- ի առկայությունից (հավելված 21-2-ում նկարագրված է apoE- ի և Ալցհայմերի հիվանդության միջև փոխհարաբերությունները):
Նկ. 21-40: Լիպոպրոտեինները և լիպիդային տրանսպորտը, և - լիպիդները տեղափոխվում են արյան մեջ լիպոպրոտեինների տեսքով, որոնք միավորված են մի շարք ֆրակցիաների ՝ տարբեր գործառույթներով և սպիտակուցների և լիպիդների տարբեր կազմերով (ներդիր 21-1, 21-2) և համապատասխանում են այդ ֆրակցիաների խտությանը: Սննդի լիպիդները համախմբվում են քիլոմիկրոնների մեջ, դրանցում պարունակվող տրիասիլգլիցերին պարունակող մեծ մասը լիպոպրոտեինային լիպազայով ազատվում են մազանոթների ոսպոզայի և մկանային հյուսվածքի մեջ: Chylomicron- ի մնացորդները (հիմնականում պարունակում են սպիտակուցներ և խոլեստերին) գրավվում են հեպատոցիտներով: Էնդոգեն լիպիդները և լյարդից խոլեստերինը հանձնվում են յուղային և մկանային հյուսվածքին ՝ VLDL ձևով: VLDL- ից լիպիդների արտազատումը (որոշ ապոլիպոպրոտեինների կորստի հետ միասին) հետզհետե վերածում է VLDLP- ի LDL- ի, ինչը խոլեստերինը փոխանցում է արտաճյուղային հյուսվածքներին կամ այն վերադարձնում է լյարդին: Լյարդը գրավում է ընկալիչներով փոխանցվող էնդոցիտոզով VLDL, LDL և քիլոմիկրոնների մնացորդները: Լրացուցիչ խոլեստերինը արտաքնային հիվանդությունների հյուսվածքներում վերափոխվում է լյարդի `LDL- ի տեսքով: Լյարդում խոլեստերինի մի մասը վերածվում է լեղու աղերի: բ - արյան պլազմայի նմուշները, որոնք վերցված են սովից (ձախ) և ճարպի բարձր պարունակությամբ սնունդ ուտելուց հետո (աջ): Fարպային սնունդ ուտելու արդյունքում ձևավորված քիլոմիկրոնները պլազմային արտաքինը նման են կաթին:

Տրիասիլգլիցերինների կորստով VLDL- ի մի մասը վերածվում է VLDL մնացորդների, որը նաև կոչվում է միջանկյալ խտության լիպոպրոտեիններ (VLDL), տրիխիլգլիցերինների հետագա հեռացումը տալիս է VLDL: ցածր խտության լիպոպրոտեիններ (LDL) (էջ 21-1): LDL ֆրակցիան, որը շատ հարուստ է խոլեստերինով և խոլեստերինի էթերերով, ինչպես նաև պարունակում է apoB-100, խոլեստերինը փոխանցում է արտագնա հյուսվածքների, որոնք իրենց պլազմային մեմբրանների վրա կրում են apoB-100 ճանաչող հատուկ ընկալիչները: Այս ընկալիչները միջնորդում են խոլեստերինի և խոլեստերինի էթերների կլանումը (ինչպես նկարագրված է ստորև):
Լրացում 21-2:ApoE ալելները որոշում են Ալցհայմերի հիվանդության դեպքը
Մարդկային բնակչության մեջ գեների կոդավորող ապոլիպոպրոտեին E.- ի երեք հայտնի տարբերակ (երեք ալել) կա ՝ APEE- ի ալելներից, APOEZ ալելը մարդու մոտ ամենատարածվածն է (մոտ 78%), APOE4 և APOE2 ալելները `համապատասխանաբար 15 և 7%: APOE4 ալելը հատկապես բնորոշ է Ալցհայմերի հիվանդությամբ տառապող մարդկանց, և այդ հարաբերությունը թույլ է տալիս կանխատեսել հիվանդության առաջացումը բարձր հավանականությամբ: Մարդիկ, ովքեր ժառանգել են APOE4- ը, մեծ Ալցհայմերի հիվանդություն զարգացնելու մեծ ռիսկ ունեն: APOE4- ի համար համասեռամոլ մարդիկ 16 անգամ ավելի մեծ հավանականությամբ են զարգացնում հիվանդությունը, հիվանդացածների միջին տարիքը մոտ 70 տարեկան է: Այն մարդկանց համար, ովքեր ժառանգում են AROEZ- ի երկու օրինակը, ընդհակառակը, Ալցհայմերի հիվանդության միջին տարիքը գերազանցում է 90 տարին:
ApoE4- ի և Ալցհայմերի հիվանդության միջև կապի մոլեկուլային հիմքը դեռ հայտնի չէ: Բացի այդ, դեռևս պարզ չէ, թե ինչպես է apoE4- ն ազդում ամիլոիդային լարերի աճի վրա, որոնք, ըստ երևույթին, Ալցհայմերի հիվանդության արմատական պատճառն են (տես Նկար 4-31, v. 1): Ենթադրությունները կենտրոնանում են apoE- ի հնարավոր դերի վրա ՝ նեյրոնների բջջային կմախքի կառուցվածքի կայունացման գործում: ApoE2 և apoEZ սպիտակուցները կապվում են մի շարք սպիտակուցների հետ, որոնք կապված են նեյրոնների միկրոթուբուլների հետ, մինչդեռ apoE4- ը չի կապում: Սա կարող է արագացնել նեյրոնների մահը: Ինչ էլ որ դառնա այդ մեխանիզմը, այս դիտարկումները հույս են ներշնչում ապոլիպոպրոտեինների կենսաբանական գործառույթների վերաբերյալ մեր պատկերացումները ընդլայնելու համար:
Լիպոպրոտեինների չորրորդ տեսակը - բարձր խտության լիպոպրոտեիններ (HDL), այս կոտորակը ձևավորվում է լյարդի և փոքր աղիքի մեջ `սպիտակուցներով հարուստ փոքր մասնիկների տեսքով, որոնք պարունակում են համեմատաբար քիչ խոլեստերին և լիովին զերծ խոլեստերինի էթերերից (Նկար 21-40): HDL- ի բաժինը պարունակում է apoA-I, apoC-I, apoC-II և այլ apolipoproteins (աղյուսակ 21-2), ինչպես նաև լեցիտին-խոլեստերին-ացիլtransferase (LC AT), ինչը կատալիզացնում է խոլեստերինի էսթերների ձևավորումը լեցիտինից (ֆոսֆատիդիլխոլին) և խոլեստերինից (Նկար 21-41): Նոր ձևավորված HDL մասնիկների մակերևույթի L CAT- ը խոլոմիկրոնի խոլեստերինի և ֆոսֆատիդիլխոլինի և VLDL մնացորդները վերածում է խոլեստերինի էթերների, որոնք սկսում են ձևավորել կորիզը ՝ նորաստեղծ դիսկոիդ HDL մասնիկները վերածելով հասուն գնդաձև HDL մասնիկների: Այս խոլեստերինով հարուստ լիպոպրոտինը վերադառնում է լյարդ, որտեղ խոլեստերինը «արտանետվում է», այս խոլեստերինից մի մասը վերածվում է աղիների աղերի:
Նկ. 21-41: Արձագանքը կատալիզացվել է լեցիտին-խոլեստերին-ացիլտրրանսֆերազով (L CAT): Այս ֆերմենտը առկա է HDL մասնիկների մակերևույթի վրա և ակտիվանում է apoA-1 (HDL ֆրակցիայի բաղադրիչ) միջոցով: Խոլեստերինի էսթերները կուտակվում են նոր ձևավորված HDL մասնիկների ներսում ՝ դրանք վերածելով հասուն HDL:

HDL- ն կարող է ներծծվել լյարդի մեջ `ընկալիչներով միջնորդավորված էնդոցիտոզով, բայց գոնե HDL խոլեստերինի որոշ չափով առնվազն որոշ մեխանիզմներով է փոխանցվում այլ հյուսվածքների: HDL մասնիկները կարող են կապվել SR - BI ընկալիչի սպիտակուցների վրա լյարդի բջիջների պլազմային մեմբրանի և ստերոիդոգեն հյուսվածքի մեջ, ինչպիսիք են երիկամային խցուկները: Այս ընկալիչները չեն միջնորդում էնդոցիտոզին, այլ խոլեստերինի և HDL ֆրակցիայի այլ լիպիդների մասնակի և ընտրովի փոխանցում բջիջ: «Plորացված» HDL- ի կոտորակը կրկին մտնում է արյան մեջ, որտեղ այն ներառում է քիլոմիկրոնների և VLDL մնացորդների լիպիդների նոր բաժիններ: Նույն HDL- ն կարող է նաև գրավել արտաճնային հյուսվածքներում պահվող խոլեստերին և այն փոխանցել լյարդին հակադարձ խոլեստերին տեղափոխումը (Նկար 21-40): Հակադարձ տրանսպորտային տարբերակներից մեկում, արդյունքում ստացված HDL- ի փոխազդեցությունը խոլեստերինով հարուստ բջիջներում գտնվող SR-BI ընկալիչների հետ, խոլեստերինի պասիվ ցրվածություն է առաջացնում բջջային մակերևույթից HDL մասնիկների մեջ, որոնք այնուհետև խոլեստերինը փոխանցում են լյարդին: Հարուստ խոլեստերինի խցում հակադարձ փոխադրման մեկ այլ տարբերակում ՝ HDL- ի պեղումից հետո, apoA-I- ը փոխազդում է ակտիվ փոխադրողի ՝ ABC սպիտակուցի հետ: ApoA-I- ը (և, ենթադրաբար, HDL) ներծծվում է էնդոցիտոզով, այնուհետև կրկին գաղտնազերծվում է, բեռնված խոլեստերինով, որը տեղափոխվում է լյարդ:
Սպիտակուցներ ABC1- ը բազմաթիվ դեղերի կրողների մեծ ընտանիքի մի մասն է, այդ կրիչները երբեմն կոչվում են ABC փոխադրողներ, քանի որ դրանք բոլորն էլ պարունակում են ATP- կապակցող ձայներիզներ (ATP - կապող ձայներիզներ), դրանք ունեն նաև երկու տրանսմբրանային տիրույթ, վեց տրանսմեբրենային խխունջներով (տե՛ս գլ. 11, v. 1): Այս սպիտակուցները ակտիվորեն փոխանցում են բազմաթիվ իոններ, ամինաթթուներ, վիտամիններ, ստերոիդ հորմոններ և աղիների աղեր ՝ պլազմային մեմբրանների միջոցով: Փոխադրողների այս ընտանիքի մեկ այլ ներկայացուցիչ է CFTR սպիտակուցը, որը, կիստիկ ֆիբրոզով, վնասվում է (տես ՝ ավելացնել 11-3, v. 1):
Խոլեստերինի էսթերը մտնում են բջիջ ընկալիչի միջնորդավորված էնդոցիտոզի միջոցով
Արյան մեջ յուրաքանչյուր LDL մասնիկ պարունակում է apoB-100, որը ճանաչվում է հատուկ մակերևութային ընկալիչների սպիտակուցներով -LDL ընկալիչները խոլեստերին գրավելու համար անհրաժեշտ բջիջների մեմբրանի վրա: LDL- ի LDL ընկալիչին կապելը առաջացնում է էնդոցիտոզ, որի պատճառով LDL- ն և դրա ընկալիչը տեղափոխվում են էնդոզոմի ներսում գտնվող բջիջ (Նկար 21-42): Էնդոզոմը ի վերջո միաձուլվում է լիզոզոմի հետ, որն իր մեջ պարունակում է ֆերմենտներ, որոնք հիդրոլիզացնում են խոլեստերինի եթերները ՝ ազատելով խոլեստերինը և ճարպաթթուները ցիտոզոլի մեջ: LDL- ից ApoB-100- ը նույնպես քայքայվում է `կազմելով ամինաթթուներ, որոնք գաղտնազերծվում են ցիտոսոլի մեջ, բայց LDL ընկալիչը խուսափում է քայքայվելուց և վերադառնում է բջիջների մակերեսին` կրկին մասնակցելու LDL- ի ներգրավմանը: ApoB-100- ը նույնպես առկա է VLDL- ում, բայց դրա ընկալիչներով կապող տիրույթը ի վիճակի չէ կապել LDL ընկալիչի հետ; VLDLP- ի LDL- ի վերափոխումը հասանելի է դարձնում ընկալիչների միացնող տիրույթը apoBB-100: Արյան խոլեստերինի փոխադրման այս ուղին և թիրախային հյուսվածքներում նրա ընկալիչներով միջնորդավորված էնդոցիտոզը ուսումնասիրվել են Մայքլ Բրաունի և Josephոզեֆ Գոլդշտեյնի կողմից:
Մայքլ Բրաուն և Josephոզեֆ Գոլդշտեյն

Նկ. 21-42: Խոլեստերին գրավելը ընկալիչներով փոխանցվող էնդոցիտոզով:

Խոլեստերինը, որն այս կերպ մտնում է բջիջները, կարող է ներառվել մեմբրանների մեջ կամ վերափոխվել է ACAT- ի (Նկար 21-38) `լիպիդային կաթիլների ներսում ցիտոզոլում պահելու համար: Երբ արյան LDL բաժնում կա բավարար խոլեստերին, ավելցուկային ներբջջային խոլեստերինի կուտակումն կանխվում է `նվազեցնելով դրա սինթեզի արագությունը:
LDL ընկալիչը նույնպես կապվում է apoE- ի հետ և զգալի դեր է խաղում լյարդի կողմից քիլոմիկրոնների և VLDL մնացորդների յուրացման մեջ: Այնուամենայնիվ, եթե LDL ընկալիչները մատչելի չեն (ինչպես, օրինակ, մկնիկի շտամում բացակայող LDL ընկալիչ գենով), VLDL մնացորդները և քիլոմիկրոնները դեռ ներծծվում են լյարդի կողմից, չնայած LDL- ն չի ներծծվում: Սա ցույց է տալիս օժանդակ պահուստային համակարգի առկայություն VLDL և քիլոմիկրոնի մնացորդների ընկալիչներով միջնորդավորված էնդոցիտոզի համար: Պահուստային ընկալիչներից մեկը LRP սպիտակուցն է (լիպոպրոտեինային ընկալիչի հետ կապված սպիտակուցը), որը կապված է լիպոպրոտեինային ընկալիչների հետ, որը կապվում է apoE- ի և մի շարք այլ լիգանների հետ:
Խոլեստերինի բիոսինթեզի կարգավորման մի քանի մակարդակ
Խոլեստերինի սինթեզը բարդ և էներգետիկորեն թանկ գործընթաց է, ուստի պարզ է, որ մարմինը ձեռնտու է ունենալ խոլեստերինի բիոսինթեզը կարգավորելու մեխանիզմ, որն իր գումարը լրացնում է ի լրումն սննդի հետ միասին: Կաթնասունների մեջ խոլեստերինի արտադրությունը կարգավորվում է ներբջջային կոնցենտրացիայի միջոցով
խոլեստերին և հորմոններ գլյուկագոն և ինսուլին: HMG - CoA- ի mevalonate- ի վերափոխման փուլը (Նկար 21-34) սահմանափակում է խոլեստերինի ձևավորման նյութափոխանակային ուղու արագությունը (կարգավորման հիմնական կետը): Այս արձագանքը կատալիզացվում է HMG - CoA reductase- ի միջոցով: Կարգավորումը ի պատասխան խոլեստերինի մակարդակի փոփոխություններին ի պատասխան միջնորդվում է վերափոխման կարգավորման էլեգանտ համակարգով `HMG - CoA ռեդուկտազը կոդավորող գենի համար: Այս գենը, ավելի քան 20 այլ գեների կոդավորող ֆերմենտների հետ միասին, որոնք ներգրավված են խոլեստերինի և չհագեցած ճարպաթթուների կլանման և սինթեզի մեջ, վերահսկվում է սպիտակուցների մի փոքր ընտանիքի կողմից, որը կոչվում է սպիտակուցներ, որոնք փոխազդում են սպիտակուցների ձևավորման ստերոլ-կարգավորող տարրի հետ (SREBP, ստերոլ կարգավորող տարրը պարտադիր սպիտակուցներ): . Սինթեզից հետո այդ սպիտակուցները ներմուծվում են էնդոպլազմիկ ցանցաթաղանթ: Միակ լուծելի ամինո տերմինալի SREBP տիրույթը գործում է որպես տառադարձման ակտիվացուցիչ ՝ օգտագործելով Ch- ում նկարագրված մեխանիզմները: 28 (v. 3): Այնուամենայնիվ, այս տիրույթը չունի միջուկ, և չի կարող մասնակցել գենի ակտիվացմանը, քանի դեռ այն մնում է SREBP մոլեկուլում: HMG գենի - CoA ռեդուկտազայի և այլ գեների արտագրությունն ակտիվացնելու համար տրանսգրացիոն ակտիվ դոմենը SREBP- ի մնացած մասերից առանձնացված է պրոտեոլիտիկ հատումով: Երբ խոլեստերինը բարձր է, SREBP սպիտակուցները անգործունակ են, ամրագրված են ER- ի վրա բարդության մեջ մեկ այլ սպիտակուցով, որը կոչվում է SCAP (SREBP - ճեղքող ակտիվացնող սպիտակուց) (Նկար 21-43): Դա SCAP- ն է, որը կապում է խոլեստերինը և մի շարք այլ ստերոլներ ՝ հանդես գալով որպես ստերոլային սենսոր: Երբ ստերոլի մակարդակը բարձր է, SCAP - SREBP համալիրը, հավանաբար, շփվում է մեկ այլ սպիտակուցի հետ, որն ամբողջ բարդությունը պահում է ER- ում: Երբ բջիջում ստերոլների մակարդակը իջնում է, SCAP- ի կոնֆորմացիոն փոփոխությունը հանգեցնում է պահպանման գործունեության կորստի, իսկ SCAP - SREBP համալիրը տեղափոխվում է վեզիկլների ներսում դեպի Գոլգի համալիր: Golgi համալիրում SREBP սպիտակուցները երկու անգամ մաքրվում են երկու տարբեր պրոտեազներով, իսկ երկրորդ ճեղքումը ՝ ազատելով ամինո-տերմինալային տիրույթը ցիտոսոլի մեջ: Այս տիրույթը տեղափոխվում է միջուկ և ակտիվացնում է թիրախային գեների տառադարձումը: SREBP սպիտակուցի ամինո-տերմինալային տիրույթը ունի կարճ կիսամյակ և արագորեն քայքայվում է պրոտեազոմներով (տես Նկար 27-48, տ. 3): Երբ ստերոլի մակարդակը բավականաչափ բարձրանում է, ամինային տերմինով SR EBP սպիտակուցային տիրույթների պրոտեոլիտիկ արտազատումը կրկին արգելափակվում է, և առկա ակտիվ տիրույթների պրոտեազոմի դեգրադացիան հանգեցնում է թիրախային գեների արագ անջատմանը:
Նկ. 21-43: SR EBP- ի ակտիվացում: SREB P սպիտակուցները, որոնք փոխազդում են ստերոլի կարգավորվող տարրի հետ (կանաչ գույն), սինթեզումից անմիջապես հետո, ներմուծվում են ER- ում ՝ կազմելով բարդույթ S SAP (կարմիր գույն): (N և C- ը նշում են սպիտակուցների ամին և կարբոքսիլ ծայրերը:) S-CAP- ի սահմանած վիճակում SRE BP սպիտակուցները անգործ են: Երբ ստերոլի մակարդակը նվազում է, SR EBP-S CAP համալիրը գաղթում է Golgi համալիր, և SR EBP սպիտակուցները հաջորդաբար մաքրվում են երկու տարբեր պրոտեազներով: Ազատագրված ամինաթթուների տերմինալի SR EBP սպիտակուցային տիրույթը գաղթում է կորիզը, որտեղ այն ակտիվացնում է ստերոլի կարգավորվող գեների արտագրությունը:

Խոլեստերինի սինթեզը կարգավորվում է նաև մի քանի այլ մեխանիզմներով (Նկար 21-44): Հորմոնալ հսկողությունը միջնորդվում է NM G-CoA ռեդուկտազի կովալենտ ձևափոխմամբ: Այս ֆերմենտը գոյություն ունի ֆոսֆորիլացված (ոչ ակտիվ) և դեֆոսֆորիլացված (ակտիվ) ձևերում: Գլյուկագոնը խթանում է ֆերմենտի ֆոսֆորիլացումը (անակտիվացումը), իսկ ինսուլինը նպաստում է dephosphorylation- ին, ակտիվացնելով ֆերմենտը և նպաստելով խոլեստերինի սինթեզին: Խոլեստերինի բարձր ներբջջային կոնցենտրացիաները ակտիվացնում են ASAT- ը, ինչը մեծացնում է խոլեստերինի էսթերիացիան նստվածքների համար: Վերջապես, բջջային խոլեստերինի բարձր մակարդակը խանգարում է գենի արտագրմանը, որը կոդավորում է LDL ընկալիչը ՝ նվազեցնելով այս ընկալիչի արտադրությունը և, հետևաբար, արյան մեջ խոլեստերինի կլանումը:
Նկ. 21-44: Խոլեստերինի մակարդակի կարգավորումը ապահովում է հավասարակշռություն սննդից խոլեստերինի սինթեզի և կլանման միջև: Գլյուկագոնը հեշտացնում է NM G -CoA ռեդուկտազի ֆոսֆորիլացումը (անգործությունը), ինսուլինը նպաստում է դեֆոսֆորիլացմանը (ակտիվացում): X - չճանաչված խոլեստերինի մետաբոլիտներ, որոնք խթանում են NM G -CoA ռեդուկտազի պրոտեոլիզը:

Չկարգավորված խոլեստերինը կարող է հանգեցնել լուրջ հիվանդության մարդկանց: Երբ սննդից ստացված սինթեզված խոլեստերինի և խոլեստերինի ընդհանուր քանակը գերազանցում է մեմբրանային հավաքման համար անհրաժեշտ քանակությունը, լեղու աղերի և ստերոիդների սինթեզը, կարող են հայտնվել արյան անոթներում խոլեստերինի պաթոլոգիական կուտակումները (աթերոսկլերոզային սալեր), ինչը հանգեցնում է դրանց արգելափակման (աթերոսկլերոզ): Արդյունաբերականացված երկրներում դա սրտի անբավարարություն է `կորոնար զարկերակների խցանումների պատճառով, ինչը մահացության հիմնական պատճառն է: Աթերոսկլերոզի զարգացումը կապված է արյան խոլեստերինի բարձր մակարդակի հետ և հատկապես LDL ֆրակցիայի հետ հանդուրժող բարձր խոլեստերինի հետ, արյան բարձր մակարդակի HDL, ընդհակառակը, բարենպաստորեն ազդում է արյան անոթների վիճակի վրա:
Ժառանգական հիպերխոլեստերեմիայի (գենետիկ թերություն) արյան խոլեստերինի մակարդակը շատ բարձր է. Ծանր մարդկանց մոտ այս տարիքում զարգանում է ծանր աթերոսկլերոզ: Թերի LDL ընկալիչի պատճառով տեղի է ունենում LDL խոլեստերին խթանող անբավարար ընկալիչի ընդունում: Արդյունքում, խոլեստերինը չի հանվում արյան հոսքից, այն կուտակում է և նպաստում է աթերոսկլերոզային սալերի ձևավորմանը: Էնդոգեն խոլեստերինի սինթեզը շարունակվում է ՝ չնայած արյան մեջ խոլեստերինի ավելցուկին, քանի որ արտաբջջային խոլեստերինը չի կարող մտնել բջիջ ՝ ներգանգային սինթեզը կարգավորելու համար (Նկար 21 -44):Ժառանգական հիպերխոլեստերեմիայի հիվանդների և շիճուկի բարձրացված խոլեստերինի հետ կապված այլ հիվանդությունների բուժման համար օգտագործվում են ստատինի դասեր: Դրանց մի մասը ստացվում է բնական աղբյուրներից, իսկ մյուսները սինթեզվում են դեղագործական արդյունաբերության կողմից: Ստատինները նման են մեվալոնատին (ավելացնել 21-3) և հանդիսանում են NMS-CoA ռեդուկտազի մրցակցային խանգարողներ:
Հավելված 21-3: Բժշկություն: Լիպիդային վարկածը և ստատինների ստեղծումը
Սրտի կորոնար հիվանդությունը (CHD) զարգացած երկրներում մահացության հիմնական պատճառն է: Կորոնարային զարկերակների նեղացումը, որոնք արյուն են բերում սրտին, տեղի է ունենում ճարպային ավանդների ձևավորման արդյունքում, որը կոչվում է աթերոսկլերոզային սալեր, այս սալերը պարունակում են խոլեստերին, ֆիբրիլային սպիտակուցներ, կալցիում, թրոմբոցների խցանումներ և բջջային բեկորներ: XX դարում: Ակտիվ քննարկումներ են եղել զարկերակային խանգարման (աթերոսկլերոզի) և արյան խոլեստերինի փոխհարաբերությունների վերաբերյալ: Այս քննարկումներն ու այս ուղղությամբ ակտիվ հետազոտությունները հանգեցրել են արդյունավետ դեղամիջոցների ստեղծմանը, որոնք իջեցնում են խոլեստերինը:
1913-ին, Ն.Ն. Անիչկովը, ռուս հայտնի գիտնական և փորձարարական պաթոլոգիայի բնագավառի մասնագետ, հրապարակեց մի աշխատանք, որում նա ցույց տվեց, որ խոլեստերինով հարուստ սնունդով սնվող նապաստակները վնաս են հասցնում արյան անոթներին, որոնք նման են տարեց մարդկանց անոթներին ՝ աթերոսկլերոզային սալերի: Անիչկովը մի քանի տասնամյակ անցկացրեց իր հետազոտությունը և արդյունքները հրապարակեց արևմտյան հայտնի ամսագրերում: Դժբախտաբար, նրա տվյալները չեն հիմք հանդիսացել մարդկանց մոտ աթերոսկլերոզի զարգացման համար մոդելի համար, քանի որ այդ ժամանակ գերակշռում էր վարկածը, որ այս հիվանդությունը ծերացման բնական արդյունք է և չի կարող կանխվել: Այնուամենայնիվ, ապացույցներն աստիճանաբար կուտակվում էին շիճուկի խոլեստերինի և աթերոսկլերոզի զարգացման (լիպիդային վարկածի) միջև փոխհարաբերությունների միջև, և 1960-ականներին: որոշ հետազոտողներ հստակ ասում են, որ այս հիվանդությունը կարելի է բուժել դեղերով: Այնուամենայնիվ, հակառակ տեսակետը գոյություն ուներ մինչև 1984 թ. Հրապարակումը ՝ ԱՄՆ Առողջապահության ազգային ինստիտուտի կողմից անցկացված խոլեստերինի դերի լայն ուսումնասիրության արդյունքների հրապարակման արդյունքների վերաբերյալ: Ուցադրվել է սրտամկանի ինֆարկտի հաճախության և արյան խոլեստերինի նվազմամբ ինսուլտի հաճախականության վիճակագրական զգալի նվազում: Այս ուսումնասիրության մեջ խոլեստերինը, անիոն փոխանակման խեժը, որը կապում է լեղու թթուները, օգտագործվել է խոլեստերին իջեցնելու համար: Արդյունքները խթանել են նոր, ավելի հզոր բուժական դեղամիջոցների որոնումը: Պետք է ասեմ, որ գիտական աշխարհում լիպիդային վարկածի վավերականության մասին կասկածներն ամբողջությամբ անհետացել են միայն ստատինների գալուստով `1980-ականների վերջին - 1990-ականների սկզբին:
Առաջին ստատինը հայտնաբերվել է Ակիրա Էնդոյի կողմից Տոկիոյի Սանկիոյում: Էնդոն հրատարակեց իր աշխատանքը 1976-ին, չնայած նա զբաղվում էր մի քանի տարի խոլեստերինի նյութափոխանակության խնդրով: 1971-ին նա առաջարկեց, որ խոլեստերինի սինթեզի խանգարիչները կարող են պարունակվել նաև այդ ժամանակ ուսումնասիրված հակաբիոտիկների սնկերի արտադրողների մեջ: Մի քանի տարվա ինտենսիվ աշխատանքի համար նա վերլուծեց տարբեր սնկերի ավելի քան 6000 մշակույթ, մինչև հասավ դրական արդյունքի: Արդյունքում առաջացող բաղադրությունը կոչվում էր կոմպակտին: Այս նյութը խոլեստերինը ցածրացրեց շների և կապիկների մեջ: Այս ուսումնասիրությունները գրավեցին Տեխասի հարավարևմտյան բժշկական դպրոցի Մայքլ Բրաունի և ofոզեֆ Գոլդշտեյնի ուշադրությունը: Բրաունը և Գոլդշտեյնը Էնդոյի հետ միասին սկսեցին համատեղ ուսումնասիրություն և հաստատեցին նրա տվյալները: Առաջին կլինիկական փորձարկումների հիմնական հաջողությունները ներգրավեցին դեղագործական ընկերությունները այս նոր դեղամիջոցների զարգացման մեջ: Merck- ում Ալֆրեդ Ալբերտսի և Ռոյ Վագելոսի գլխավորությամբ մի խումբ նախաձեռնել է սնկային մշակույթների նոր ցուցադրություն և, ընդհանուր առմամբ 18 մշակույթների վերլուծության արդյունքում, հայտնաբերել է ևս մեկ ակտիվ դեղամիջոց: Նոր նյութը կոչվում է լովաստատին: Այնուամենայնիվ, միևնույն ժամանակ, լայնորեն հավատում էին, որ կոմպակտինի բարձր չափաբաժինների օգտագործումը շներին հանգեցնում է քաղցկեղի զարգացմանը և նոր ստատինների որոնում 1980-ականներին: կասեցվել է: Այնուամենայնիվ, այդ ժամանակ արդեն ակնհայտ էին ստատինների օգտագործման առավելությունները ընտանեկան հիպերխոլեստերեմիայի հիվանդների բուժման համար: Միջազգային փորձագետների և Պարենի և դեղերի վարչության (FDA, ԱՄՆ) բազմաթիվ խորհրդակցություններից հետո Մերքը սկսեց զարգացնել լովաստատինը: Հաջորդ երկու տասնամյակների լայնածավալ ուսումնասիրությունները չեն պարզել լովաստատինի քաղցկեղածին ազդեցությունը և նոր սերնդի դեղեր, որոնք հայտնվել են դրանից հետո:
Նկ. 1. Ստատինները NM G-CoA ռեդուկտազի խանգարիչ են: Մեվալոնատի և չորս դեղագործական արտադրանքի (ստատինների) կառուցվածքի համեմատություն, որոնք խանգարում են NM G -CoA ռեդուկտազի գործողությունը:

Ստատիններ արգելակել HMG - CoA- ի գործողությունը `ռեդուկտազը, ընդօրինակելով mevalonate- ի կառուցվածքը, և դրանով իսկ արգելափակել խոլեստերինի սինթեզը: LDL ընկալիչի գենի մեկ օրինակի թերության հետևանքով առաջացած հիպերխոլեստերեմիայի հիվանդների մոտ, լովաստատին ընդունելիս, խոլեստերինի մակարդակը կրճատվում է 30% -ով: Դեղը նույնիսկ ավելի արդյունավետ է զուգակցված հատուկ խեժերի հետ, որոնք կապում են աղիների թթուները և կանխում են դրանց հակադարձ կլանումը աղիքներից:
Ներկայումս ստատիններն առավել հաճախ օգտագործվում են արյան պլազմային խոլեստերինը իջեցնելու համար: Medicationանկացած դեղորայք ընդունելիս հարց է ծագում դրանց անցանկալի կողմնակի բարդությունների մասին: Այնուամենայնիվ, ստատինների դեպքում շատ կողմնակի բարդություններ, հակառակը, դրական են: Այս դեղերը կարող են խթանել արյան հոսքը, շտկել արդեն իսկ առկա աթերոսկլերոզային սալերը (այնպես, որ դրանք չփչանան արյան անոթների պատերից և չխանգարեն արյան հոսքին), խանգարել թրոմբոցիտների ագրեգացումը, ինչպես նաև թուլացնել արյան անոթների պատերին բորբոքային պրոցեսները: Առաջին անգամ ստատիններ ընդունող հիվանդների դեպքում այդ հետևանքները դրսևորվում են նույնիսկ նախքան խոլեստերինի մակարդակի նվազումը, և, հավանաբար, կապված են իզոպրենոիդ սինթեզի արգելքի հետ: Իհարկե, ստատինների ոչ բոլոր կողմնակի ազդեցությունները ձեռնտու չեն: Որոշ հիվանդների մոտ (սովորաբար ստատիններ ընդունող հիվանդների հետ միասին, որոնք իջեցնում են խոլեստերինը), մկանների ցավը և մկանների թուլությունը կարող են առաջանալ, իսկ երբեմն էլ ՝ բավականին ուժեղ ձևով: Գրանցված են նաև ստատինների այլ բավականին շատ կողմնակի բարդություններ, որոնք, բարեբախտաբար, հազվադեպ են լինում: Հիվանդների ճնշող մեծամասնության դեպքում ստատիններ վերցնելը կարող է խանգարել սրտանոթային հիվանդությունների զարգացումը: Anyանկացած այլ դեղորայքի նման, ստատինները պետք է օգտագործվեն միայն ձեր բժշկի առաջարկած համաձայն:

HDL խոլեստերինի ժառանգական բացակայությամբ խոլեստերինի մակարդակը շատ ցածր է, Tangier հիվանդության դեպքում խոլեստերինը գործնականում որոշված չէ: Երկու գենետիկական խանգարումները առաջանում են ABC1 սպիտակուցի մուտացիաներից: HDL- ի խոլեստերինի կոլեկտորը չի կարող խոլեստերինը գրավել ABC1- ի անբավարար բջիջներից, իսկ խոլեստերինով քայքայված բջիջները արագորեն հանվում են արյունից և ոչնչացվում: Թե HDL- ի, թե Tangier հիվանդության ժառանգական բացակայությունը շատ հազվադեպ են (ամբողջ աշխարհում հայտնի են Tangier հիվանդությամբ 100-ից ավելի ընտանիքներ), բայց այդ հիվանդությունները ցույց են տալիս ABC1 սպիտակուցի դերը HDL պլազմայի մակարդակը կարգավորելու հարցում: Քանի որ պլազմային HDL- ի ցածր մակարդակը կապված է կորոնար շնչերակներից վնասվածքի բարձր տեմպի հետ, ABC1 սպիտակուցը կարող է օգտակար թիրախ լինել այն դեղերի համար, որոնք նախատեսված են HDL- ի մակարդակները կարգավորելու համար: ■
Ստերոիդ հորմոնները ձևավորվում են խոլեստերինի կողային շղթան բաժանելու և դրա օքսիդացման միջոցով:
Մարդը ստանում է իր ստերոիդային բոլոր հորմոնները խոլեստերինից (Նկար 21-45): Երկու ստերոիդ հորմոններ սինթեզվում են մակերիկամային ծառի կեղեվում. հանքային կորտիկոիդներ,որոնք կարգավորում են անօրգանական իոնների (Na +, C լ և HC O) կլանումը 3 -) երիկամներում, և գլյուկոկորտիկոիդներ, որոնք օգնում են կարգավորել գլյուկոնեոգենեզը և նվազեցնել բորբոքային պատասխանը: Սեռական հորմոնները արտադրվում են տղամարդկանց և կանանց վերարտադրողական բջիջներում և պլասենցայում: Նրանց մեջ պրոգեստերոն որը կարգավորում է կանանց վերարտադրողական ցիկլը, androgens (օրինակ, տեստոստերոն) և էստրոգեններ (էստրադիոլ), որոնք ազդում են համապատասխանաբար տղամարդկանց և կանանց երկրորդական սեռական բնութագրերի զարգացման վրա: Ստերոիդ հորմոնները ազդեցություն ունեն շատ ցածր կոնցենտրացիաների մեջ, ուստի սինթեզվում են համեմատաբար փոքր քանակությամբ: Լեղու աղերի հետ համեմատաբար, քիչ խոլեստերին է սպառում ստերոիդ հորմոնների արտադրության համար:

Նկ. 21-45: Խոլեստերինից ձևավորվում են որոշ ստերոիդ հորմոններ: Այս միացություններից մի քանիսի կառուցվածքները ներկայացված են Նկ. 10-19, v. 1:

Ստերոիդ հորմոնների սինթեզը պահանջում է ածխածնի մի քանի կամ բոլոր ատոմների հեռացում խոլեստերինի C-17 D- օղակի «կողմնակի շղթայում»: Կողմնակի շղթայի հեռացումը տեղի է ունենում ստերոիդոգեն հյուսվածքների mitochondria- ում: Հեռացման գործընթացը բաղկացած է կողմնակի շղթայի երկու հարակից ածխածնի ատոմների հիդրօքսիլացումից (C-20 և C-22), այնուհետև դրանց միջև կապի պեղումը (Նկար 21-46): Տարբեր հորմոնների ձևավորումը ներառում է նաև թթվածնի ատոմների ներդրում: Ստերոիդ բիոսինթեզի ընթացքում հիդրօքսիլացման և օքսիդացման բոլոր ռեակցիաները կատալիզացվում են խառը գործառույթով օքսիդազներով (ավելացնել 21-1), որոնք օգտագործում են NА D PH, O 2 և միտոքոնդրիալ ցիտոքրոմ P-450:
Նկ. 21-46: Կողմնակի շղթայի մանրացումը ստերոիդ հորմոնների սինթեզում: Այս օքսիդազային համակարգում խառը գործառույթով, որը օքսիդացնում է հարակից ածխածնի ատոմները, ցիտոխրոմ P-450- ը հանդես է գալիս որպես էլեկտրոնային կրիչ: Գործընթացին մասնակցում են նաև էլեկտրոն տեղափոխող սպիտակուցներ, adrenodoxin և adrenodoxin reductase: Կողնային շղթայի բաժանման այս համակարգը հայտնաբերվել է վերերիկամային ծառի կեղեվակի mitochondria- ում, որտեղ տեղի է ունենում ստերոիդների ակտիվ արտադրություն: Pregnenolone- ը բոլոր մյուս ստերոիդ հորմոնների նախադրյալն է (Նկար 21-45):

Խոլեստերին բիոսինթեզի միջնորդները ներգրավված են շատ այլ նյութափոխանակության ուղիներ:
Ի լրումն իր դերի ՝ որպես խոլեստերինի բիոսինթեզի միջնորդ, իզոպանտենիլ պիրոֆոսֆատը ծառայում է որպես ակտիվացված նախադրյալ `կենսաբանական տարբեր ֆունկցիաներ իրականացնող հսկայական քանակի բիոմոլեկուլների սինթեզում (Նկար 21-47): Դրանք պարունակում են A, E և K վիտամիններ, բույսերի պիգմենտներ, ինչպիսիք են կարոտինը և քլորոֆիլ ֆիտոլի շղթան, բնական ռետինները, շատ եթերայուղերը (օրինակ ՝ կիտրոնի յուղի բուրավետ հիմքը, էվկալիպտը, մուշկը), միջատների անչափահաս հորմոնը, որը կարգավորում է մետամորֆոզը, դոլիչոլները: ծառայում են որպես լիպիդ լուծվող կրողներ `պոլիսաքարիդների, ubiquinone- ի և plastoquinone- ի բարդ սինթեզում` էլեկտրոնային կրիչներ `mitochondria- ի և chlorlorasts- ի մեջ: Այս բոլոր մոլեկուլները կառուցվածքում իզոպրենոիդներ են: Բնության մեջ հայտնաբերվել են ավելի քան 20,000 տարբեր իզոպրենոիդներ, և ամեն տարի հարյուրավոր նորեր են գրանցվում:
Նկ. 21-47: Իզոպրենոիդների բիոսինթեզի ընդհանուր պատկերը: Այստեղ ներկայացված վերջնական արտադրանքի մեծ մասի կառուցվածքները ներկայացված են բաժնում: 10 (v. 1):

Պրենիլացումը (իզոպրենոիդների կովալենտային կցորդ, տես Նկար 27-35) ընդհանուր մեխանիզմ է, որի միջոցով սպիտակուցները խարսխվում են կաթնասունների բջիջների մեմբրանների ներքին մակերեսի վրա (տես Նկար 11-14): Որոշ սպիտակուցներում պարտավորված լիպիդը ներկայացված է 15-ածխածնային ֆարնեսիլ խմբի կողմից, մյուսներում `20-ածխածնի geranyl geranyl խումբ: Այս երկու տեսակի լիպիդները կցում են տարբեր ֆերմենտներ: Հնարավոր է, որ նախածննդյան ռեակցիաները սպիտակուցներն ուղղում են տարբեր մեմբրաններ, կախված նրանից, թե որ լիպիդն է կցված: Սպիտակուցի պարարտացումը ևս մեկ կարևոր դեր է իզոպրեն ածանցյալների համար `խոլեստերինի նյութափոխանակության ուղու մասնակիցները:
21.4 Բաժնի ամփոփ նկարագիրը Խոլեստերինի, ստերոիդների և իզոպրենոիդների կենսոսինթեզ
■ Խոլեստերինը ձևավորվում է ացետիլ-CoA- ից `ռեակցիայի բարդ հաջորդականությամբ միջնորդների միջոցով, ինչպիսիք են β-հիդրօքսի-β-մեթիլգլուտարիլ-CoA- ն, մեվալոնատը, երկու ակտիվացված isoprene dimethylallyl պիրոֆոսֆատը և isopentenyl պիրոֆոսֆատը: Իզոպրենի ստորաբաժանումների խտացումը տալիս է ոչ ցիկլային սկկալին, որը ցիկլիզացնում է `կազմելով խտացրած օղակաձև համակարգ և ստերոիդային կողմնային ցանց:
Of Խոլեստերինի սինթեզը գտնվում է հորմոնալ հսկողության տակ և, բացի այդ, խանգարվում է ներբջջային խոլեստերինի կոնցենտրացիաների մեծացմանը, ինչը տեղի է ունենում կովալենտ ձևափոխման և փոխպատվաստման կարգավորման միջոցով:
■ Խոլեստերինի և խոլեստերինի էսթերներն արյան մեջ փոխանցվում են որպես պլազմային լիպոպրոտեիններ: VLDL մասնաբաժինը խոլեստերինը, խոլեստերինի էթերները և տրիասիլգլիցերիները փոխանցում է լյարդից այլ հյուսվածքներ, որտեղ տրիխիլգլիցերինները մաքրվում են լիպոպրոտեինային լիպազով, իսկ VLDL- ը վերածվում է LDL: Խոլեստերինի և խոլեստերինի էսթերներում հարստացված LDL հատվածը անուղղակիորեն գրավվում է ընկալիչների կողմից էնդոցիտոզով, մինչդեռ LDL- ում B-100 apolipoprotein- ը ճանաչվում է պլազմային թաղանթային ընկալիչների կողմից: HDL- ն արյան մեջ հեռացնում է խոլեստերինը ՝ այն փոխանցելով լյարդին: Խոլեստերինի նյութափոխանակության մեջ սննդային պայմանները կամ գենետիկական արատները կարող են հանգեցնել աթերոսկլերոզի և սրտամկանի ինֆարկտի:
■ Ստերոիդ հորմոնները (գլյուկոկորտիկոիդներ, միներալոկորտիկոիդներ և սեռական հորմոններ) ձևավորվում են խոլեստերինից ՝ փոխելով կողմնային շղթան և օղակների թթվածինները ներմուծելով օղակների ստերոիդ համակարգում: Շատ այլ isoprenoid միացություններ արտադրվում են mevalonate- ից `խոլեստերինի հետ մեկտեղ իզոպանտենիլ պիրոֆոսֆատի և dimethylallyl պիրոֆոսֆատի խտացումով:
Certain Որոշ սպիտակուցների նախրիլացիան նրանց ուղղորդում է բջջային մեմբրաններով կապող վայրեր և կարևոր է նրանց կենսաբանական գործունեության համար:
Հարց 48. Բարձր ճարպաթթուների նյութափոխանակության կարգավորումը (β-օքսիդացում և կենսոսինթեզ): Malonyl CoA- ի սինթեզ: Acetyl CoA carboxylase, նրա գործունեության կարգավորում: Acyl Co-a- ի փոխադրում mitochondria- ի ներքին թաղանթով:
Հիմնական
սպառվում է ֆենիլալանինի քանակը
2 եղանակով.
միացնում է
սկյուռներում,
շրջվում է
տիրոսինի մեջ:
Շրջվելով
առաջին հերթին ֆենիլալանինը `տիրոզինին
անհրաժեշտ է հեռացնել ավելցուկը
ֆենիլալանինը, քանի որ բարձր կոնցենտրացիաները
Դա բջիջների համար թունավոր է: Կրթություն
տիրոսինը իրականում նշանակություն չունի
այս ամինաթթվի պակասից ի վեր
բջիջներում գործնականում չի լինում:
Հիմնական
սկսվում է ֆենիլալանինի նյութափոխանակությունը
իր հիդրօքսիլացմամբ (Նկար 9-29), in
արդյունքում տիրոսինը:
Այս արձագանքը կատալիզացվում է հատուկի կողմից
monooxy-nase - ֆենիլալանինի հիդրա (zsilase,
որը ծառայում է որպես համահեղինակ
tetrahydrobiopterin (N4BP):
Կախված է նաև ֆերմենտային գործունեությունը
Fe2- ի առկայությունը:
Ներ
լյարդը հիմնականում արագացված մոբիլիզացումն է
գլիկոգեն (տես 7-րդ բաժինը): Այնուամենայնիվ, բաժնետոմսերը
գլիկոգենը լյարդում ավելանում է
18-24 ժամ ծոմ պահելը: Հիմնական աղբյուր
գլյուկոզա, քանի որ բաժնետոմսերը սպառվում են
գլիկոգենը դառնում է գլյուկոնեոգենեզ,
որը սկսում է արագանալ միջոցով
Նկ.
11-29: Հիմնական նյութափոխանակությունը
էներգիա, երբ ներծծողը փոխվում է
հետմահաց պետություն: ԱԹ
- ketone մարմիններ, FA - ճարպաթթուներ:
4-6 ժամ
վերջին կերակուրից հետո: Ենթավազաններ
գլիցերինը օգտագործվում է գլյուկոզայի սինթեզի համար,
ամինաթթուներ և լակտատ: Բարձր
գլյուկագոնի համակենտրոնացման սինթեզի մակարդակը
ճարպաթթուները նվազել են `պայմանավորված
ֆոսֆորիլացում և անգործություն
ացետիլ CoA կարբոքսիլազ և փոխարժեք
p- օքսիդացումը մեծանում է: Սակայն,
ավելացել է ճարպի մատակարարումը լյարդին
թթուներ, որոնք տեղափոխվում են
ճարպային պահեստներից: Ձևավորվեց ացետիլ-CoA
ճարպաթթուների օքսիդացման մեջ այն օգտագործվում է
լյարդում `ketone մարմինների սինթեզի համար:
Ներ
ճարպային հյուսվածքներ աճող համակենտրոնացումով
գլյուկագոն իջեցրեց սինթեզի արագությունը
Խթանում է TAG- ը և լիպոլիզը: Խթանում
լիպոլիզ - ակտիվացման արդյունք
հորմոնալ զգայուն TAG լիպազա
adipocytes գլյուկագոնի ազդեցության տակ:
Attyարպաթթուները դառնում են կարևոր
էներգիայի աղբյուրները լյարդի, մկանների և
ճարպային հյուսվածք:
Այսպիսով
այսպիսով, հետաբաշխման շրջանում
արյան գլյուկոզի կոնցենտրացիան պահպանվում է
80-100 մգ / դլ մակարդակով, և ճարպի մակարդակում
թթուները և ketone մարմինները մեծանում են:
Շաքարավազ
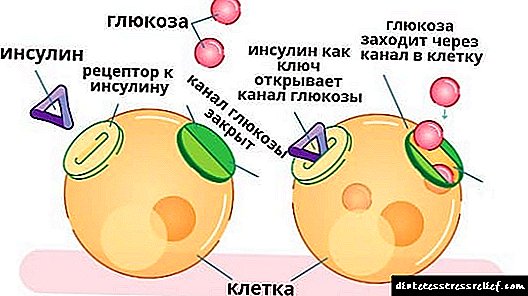
շաքարախտը հիվանդություն է, որը առաջանում է
բացարձակ կամ հարաբերական պատճառով
ինսուլինի անբավարարություն:
Ա.
Շաքարի հիմնական կլինիկական ձևերը
շաքարային դիաբետ
Համաձայն
Համաշխարհային կազմակերպություն
առողջապահական շաքարախտ
դասակարգված ըստ տարբերությունների
գենետիկ գործոնները և կլինիկական
երկու հիմնական ձև ՝ շաքարախտ
I տիպ - ինսուլին կախված (IDDM) և շաքարախտ
II տիպ - ոչ ինսուլինի անկախ (NIDDM):
Կարգավորումը
zhk- ի սինթեզը կարգավորող ֆերմենտ
LCD- ի սինթեզ - ացետիլ CoA կարբոքսիլազ:
Այս ֆերմենտը կարգավորվում է մի քանիսի միջոցով
եղանակներ
Ակտիվացում / տարաձայնություն
ֆերմենտային ենթամոնտաժային համալիրներ: Ներ
ացետիլ CoA կարբոքսիլազի անգործուն ձև
ներկայացնում է առանձին բարդույթներ,
որոնցից յուրաքանչյուրը բաղկացած է 4 ստորաբաժանումից:
Ֆերմենտի ակտիվացումը ցիտրատն է: Այն խթանում է
բարդությունների համադրություն, որպես արդյունք
որի միջոցով ֆերմենտային ակտիվությունը մեծանում է
. Inhibitor-palmitoyl-CoA: Նա զանգում է
բարդ տարանջատում և նվազում
ֆերմենտային գործունեություն:
Ֆոսֆորիլացիա / դեֆոսֆորիլացիա
ացետիլ CoA կարբոքսիլազ: Ներ
postabsorption վիճակը կամ ներսում
ֆիզիկական աշխատանքը գլյուկագոնիզացված
ադրենալին `ադենիլատիկ ցիկլազի միջոցով
համակարգը ակտիվանում է prokinase A- ի և
խթանել ենթախմբի ֆոսֆորիլացումը
ացետիլ CoA կարբոքսիլազ: Ֆոսֆորիլացված
ֆերմենտը ոչ ակտիվ է և ճարպի սինթեզը
թթուները դադարում են:
Ներծծող
ժամանակահատվածը ինսուլինն ակտիվացնում է ֆոսֆատազը,
և ացետիլ-CoA կարբոքսիլազը մտնում է
dephosphorylated պետություն: Հետո
տեղի է ունենում ցիտրատի ազդեցության տակ
ֆերմենտի պրոտոմերների պոլիմերացում, և
նա ակտիվանում է: Բացի ակտիվացումից
ֆերմենտ, ցիտրատը կատարում է մեկ այլ
գործառույթը LCD- ի սինթեզում: Ներծծող
ժամանակահատվածը լյարդի բջիջների mitochondria- ում
կուտակում է ցիտրատ, որի մեջ
տեղափոխվում է ացիլային մնացորդ
ցիտոսոլ:
Կարգավորումը
β- օքսիդացման տեմպերը:
Β- օքսիդացում-նյութափոխանակության ուղի,
ամուր կապված է CPE- ի և ընդհանուրի աշխատանքի հետ
կատաբոլիզմի եղանակներ: Հետևաբար դրա արագությունը
կարգավորվում է բջջային անհրաժեշտության դեպքում
էներգիա, այսինքն ATP / ADP և NADH / NAD հարաբերակցությամբ, ինչպես նաև CPE- ի և
կատաբոլիզմի ընդհանուր ուղին: Արագություն
Հյուսվածքների մեջ β- օքսիդացումը կախված է առկայությունից
substrate, այսինքն.
ճարպի քանակի վրա
թթուներ, որոնք մտնում են mitochondria:
Ազատ ճարպաթթուների խտացում
արյան մեջ ակտիվանում է
ծոմ պահելու ընթացքում ճարպային հյուսվածքում լիպոլիզ
գլյուկագոնի ազդեցության տակ և ֆիզիկական ընթացքում
աշխատել ադրենալինի ազդեցության տակ: Դրանց մեջ
ճարպաթթուները դառնում են
էներգիայի գերակշռող աղբյուր
մկանների և լյարդի համար `որպես արդյունք
β- օքսիդացումները ձևավորվում են NADH- ի և ացետիլ-CoA- ի խանգարմամբ
պիրուվատի dehydrogenase համալիր:
Պիրուվատի ձևավորման փոխակերպում
գլյուկոզայից մինչև ացետիլ-CoA դանդաղում է:
Միջին նյութափոխանակիչները կուտակվում են
գլիկոլիզը և, մասնավորապես, գլյուկոզա-6-ֆոսֆատը:
Գլյուկոզա-6-ֆոսֆատը խանգարում է հեքսոկինազին
և, հետևաբար, հուսահատվում է
գործընթացում գլյուկոզի օգտագործումը
գլիկոլիզ: Հետևաբար գերակշռող
LCD- ի օգտագործումը որպես հիմնական աղբյուր
էներգիան մկանային հյուսվածքում և լյարդում
խնայում է գլյուկոզան նյարդային հյուսվածքի համար և
կարմիր արյան բջիջները:
Բ-օքսիդացման մակարդակը նույնպես
կախված է ֆերմենտային գործունեությունից
carnitine acyltransferases I.
Լյարդում այս ֆերմենտը խանգարվում է:
malonyl CoA, ձևավորված նյութ
LCD- ի կենսասինթեզով: Ներծծվող ժամանակահատվածում
գլիկոլիզացումը ակտիվանում է լյարդում և
ավելանում է ացետիլ-CoA- ի ձևավորումը
պիրուվատից: Սինթեզի առաջին ռեակցիան
ացետիլ-CoA- ի LCD փոխարկումը malonyl-CoA:
Malonyl-CoA- ն խանգարում է LCD- ի β- օքսիդացմանը,
որոնք կարող են օգտագործվել սինթեզի համար
ճարպ
Կրթություն
malonyl-CoA- ը ացետիլ-CoA- կարգավորիչից
ռեակցիա կենսոսինթեզում Առաջին արձագանքը
ացետիլ-CoA- ի սինթեզի վերափոխումը դեպի
malonyl CoA: Կատալիտիկ ֆերմենտ
այս ռեակցիան (ացետիլ Coa carboxylase),
պատկանում են լիգազների դասին: Նա պարունակում է
կովալալորեն կապված բիոտին: Առաջինում
co2 կովալենտ ռեակցիայի փուլերը
էներգիայի պատճառով կապվում է բիոտինի հետ
ATP, 2-րդ փուլում COO- փոխանցված
acetyl-CoA- ի վրա `malonyl-CoA ձևավորելու համար:
Acetyl CoA Carboxylase enzyme գործունեության
որոշում է հետագա բոլորի արագությունը
սինթեզի ռեակցիաներ lc
ցիտրատը ակտիվացնում է ֆերմենտը ցիտոսոլում
ացետիլ CoA կարբոքսիլազ: Malonyl CoA in
իր հերթին խանգարում է ավելի բարձր փոխանցմանը
ճարպաթթուները ցիտոզոլից մինչև մատրիցա
mitochondria խանգարող գործունեություն
արտաքին ացետիլ CoA` carnitine acyltransferase,
այդպիսով անջատելով ավելի բարձր օքսիդացումը
ճարպաթթուներ:
Acetyl-CoA Oxaloacetate
HS-CoA ցիտրատ

HSCOA ATP Citrate → Acetyl-CoA ADP Pi Oxaloacetate
Acetyl-CoA
ցիտոպլազմում ծառայում է որպես ելակետ
lcd- ի և oxaloacetate- ի սինթեզի համար
ցիտոսոլը անցնում է վերափոխումներ
արդյունքը, որի ձևավորվում է պիրուվատը:
Խոլեստերինի բիոսինթեզ
Խոլեստերինի բիոսինթեզը տեղի է ունենում էնդոպլազմիկ ցանցաթաղանթում: Մոլեկուլում ածխածնի բոլոր ատոմների աղբյուրը ացետիլ-SCoA- ն է, որն այստեղ գալիս է mitochondria- ից `որպես ցիտրատի մի մաս, ինչպես ճարպաթթուների սինթեզում: Խոլեստերինի կենսոսինթեզը սպառում է 18 ATP մոլեկուլ և 13 NADPH մոլեկուլ:
Խոլեստերինի ձևավորումը տեղի է ունենում ավելի քան 30 ռեակցիա, որը կարող է խմբավորվել մի քանի փուլով:
1. Մեվալոնաթթվի սինթեզ:
Սինթեզի առաջին երկու ռեակցիաները համընկնում են ketogenesis- ի ռեակցիաներին, բայց 3-հիդրօքսի-3-մեթիլգլուտարիլ-ScoA- ի սինթեզից հետո ֆերմենտը մտնում է հիդրօքսիմեթիլ-գլուտարիլ-ScoA ռեդուկտազը (HMG-SCOA reductase) ՝ ձևավորելով մեվալոնաթթու:
 |